Одна из человеческих полимераз способна проводить обратную транскрипцию

Нуклеиновые кислоты — носители генетической информации у всех обитателей Земли — представлены молекулами ДНК и РНК. Принцип комплементарности позволяет строить новые цепочки нуклеиновых кислот на матрице уже существующих. В клетках эукариот есть ферменты, которые катализируют синтез цепочек ДНК на матрице ДНК (репликацию) или синтез РНК на матрице ДНК (транскрипцию). У некоторых вирусов, например у ВИЧ, имеются ферменты, катализирующие синтез цепочек ДНК на матрице РНК (обратную транскрипцию). Группа американских ученых выяснила, что такой же способностью обладает и один из ферментов наших собственных клеток — ДНК-полимераза тета. Ранее было известно, что полимераза тета, играющая важную роль в репарации повреждений ДНК, при репликации делает большое количество ошибок, вставляя некомплементарные нуклеотиды. Теперь же выяснилось, что этот фермент охотно строит ДНК на матрице РНК, и к тому же ошибки в ходе такого синтеза появляются гораздо реже. По скорости синтеза ДНК на матрице РНК и по частоте ошибок при таком синтезе полимераза тета оказалась сопоставима с обратной транскриптазой ВИЧ.
В живых клетках ДНК — молекула генетической памяти — постоянно подвергается риску появления разнообразных повреждений. Само собой, это означает необходимость регулярного обслуживания по ремонту молекулы ДНК — этот процесс называется репарацией. Для репарации клетки располагают обширным набором механизмов и ферментов — выбор определяется типом повреждений, их количеством, стадией клеточного цикла и еще рядом факторов. В 2015 году за изучение систем репарации ДНК была присуждена Нобелевская премия (см. новость Нобелевская премия по химии — 2015, «Элементы», 09.10.2015).
Полимераза тета (DNA polymerase theta, Polθ) — один из ферментов системы репарации, работающий в клетках человека и других млекопитающих. Его назначение — спасти клетку от гибели в тех условиях, когда повреждений очень много и другие системы репарации не справляются (в первую очередь это касается повреждений типа 2-цепочечных разрывов ДНК). В наших клетках эта полимераза кодируется геном POLQ. Давно известная особенность Polθ — очень низкая точность синтеза ДНК. Именно эта особенность и позволяет ему беспрепятственно пройти через «трудные» места в ДНК, содержащие множество повреждений. Более точно работающие полимеразы в таких местах просто останавливаются, не будучи способными вставить какой-то нуклеотид напротив места, содержащего повреждение. Однако за использование фермента с низкой точностью становится приходится платить — в ДНК появляется много мутаций (главным образом замен) в нуклеотидной последовательности.
Интерес ученых к Polθ обусловлен в значительной мере тем фактом, что эта полимераза гораздо активнее работает в раковых клетках, чем в здоровых клетках организма. И это одна из причин, по которой раковые клетки очень быстро мутируют и «эволюционируют» внутри организма, приобретая выгодные для себя мутации (например, делающие их более устойчивыми к воздействию противораковых лекарств). Ингибирование этого фермента даже рассматривают как одну из перспективных стратегий борьбы с проблемой развития лекарственной устойчивости в ходе лечения онкологических пациентов (T. Kent et al., 2016. DNA polymerase θ specializes in incorporating synthetic expanded-size (xDNA) nucleotides).
Эволюционно Polθ происходит от Pol I — фермента, имеющегося у большинства прокариот, который обычно обеспечивает очень точную репликацию ДНК. В структуре Polθ, как и в структуре Pol I, имеется собственно каталитический домен и домен «самоконтроля» (proofreading), который позволяет высокоточной полимеразе, в случае вставки ошибочного нуклеотида, остановиться, вырезать неправильный нуклеотид позади себя и заменить его на другой. Благодаря этому домену полимераза не двинется дальше, пока не будет встроен верный нуклеотид. Однако в Polθ этот домен в ходе эволюции оказался подпорчен несколькими мутациями, из-за чего он перестал обеспечивать такой самоконтроль.
ДНК-полимеразы бывают разные. Однако все их можно подразделить на две большие группы — в зависимости от типа субстрата. Существуют ДНК-зависимые ДНК-полимеразы — таковыми являются все ранее известные ферменты, задействованные в репликации и репарации у клеточных организмов. Из названия следует, что эти ферменты строят новые цепочки ДНК на матрице родительских цепочек ДНК. Другую группу составляют РНК-зависимые ДНК-полимеразы (их же часто называют обратными транскриптазами) — они строят цепочки ДНК, используя в качестве матриц цепочки РНК. Такие ферменты есть у некоторых вирусов и у некоторых транспозонов — геномных паразитов, имеющих, предположительно, вирусное происхождение. Вопрос о том, выполняют ли обратные транскриптазы транспозонов какие-либо полезные функции для клетки является предметом дискуссий и мы его здесь касаться не будем, потому что нас в данном случае интересует совершенно родной белок клеток животных, не имеющий признаков вирусного происхождения. Однако кое в чем он оказался схож с полимеразами вирусов, поломав такую стройную картину классификации ДНК-полимераз по субстрату, которую мы только что представили.
Главное отличие между РНК и ДНК состоит в том, что в качестве сахара — обязательного компонента нуклеиновых кислот — в первом случае используется рибоза, во втором — дезоксирибоза. В сущности, вся разница сводится к наличию или отсутствию в молекуле сахара одного атома кислорода. Однако эта разница достаточно существенна, чтобы обеспечивать ферментам высокую специфичность при выборе субстрата при осуществлении матричного синтеза ДНК — это было показано для всех ранее изучавшихся ДНК-полимераз.
Наличие неактивного домена самоконтроля натолкнуло ученых из США на гипотезу, что, возможно, Polθ способна синтезировать ДНК не только на матрице ДНК, но и на матрице РНК. Для проверки этой гипотезы и было затеяно исследование, о котором пойдет речь. Параллельные испытания провели для обратной транскриптазы ВИЧ (синтез ДНК на матрице РНК — основная профессия обратной транскриптазы) и для еще одного человеческого фермента репарации, также склонного к ошибкам, — полимеразе эта (Polη).
В первом испытании ферментам дали наращивать цепочку ДНК на матрице РНК длиной 22 нуклеотида, с которой был связан ДНК-праймер длиной 11 нуклеотидов. Результаты можно видеть на рис. 2. Пятнышки, которые мы видим на фотографии электрофорезной пластинки — это фрагменты нарощенной ДНК. Если к праймеру присоединился один нуклеотид — получаем фрагмент длиной 12, нуклеотидов, если два — длиной 13 и т. д.

Рис. 2. Оценка способности разных полимераз наращивать цепочку ДНК по матрице РНК. А — накопление продуктов реакции полимеризации с течением времени. Показаны фотографии гелей, в которых электрофорезом разделялись продукты реакции. Левое фото — при посредстве обратной транскриптазы ВИЧ HIV RT, среднее — при посредстве полимеразы тета Polθ, правое — при посредстве полимеразы эта Polη. Над фотографиями схематично изображен начальный субстрат — дуплекс из РНК-матрицы (цепочка РНК показана красным цветом) и ДНК-затравки (цепочка ДНК показана черным цветом). Затравка имеет длину 11 нуклеотидов, так что пятна в геле, напротив которых стоит отметка 11 nt — это оставшиеся не удлиненными затравки. Цепочка матрицы имеет длину 22 нуклеотида — и это теоретически максимальная возможная длина продукта матричного синтеза ДНК (в геле видны и продукты несколько большей длины — это следствие способности полимераз добавлять нуклеотид нематричным способом). Цепочки промежуточной длины — это продукты незавершенной реакции полимеризации. Можно убедиться, что эффективный синтез ДНК идет только при работе ферментов HIV RT и Polθ. B доля затравок, удлиненных в течение указанного времени при работе HIV RT и Polθ. Видно, что динамика работы этих двух ферментов очень схожа. Рисунок из обсуждаемой статьи в Science Advances
Как видно, чем больше времени давалось ферменту (до 32 минут в общей сложности), тем больше получалось длинных фрагментов (в этой системе максимально возможными были фрагменты длиной 22 нуклеотида) — причем, только если в качестве фермента выступали Polθ или обратная транскриптаза ВИЧ (HIV RT). А вот фермент Polη в тех же концентрациях и за то же время успевал добавить не больше трех (правда при очень сильном повышении концентрации этого фермента удавалось добиться получения более протяженных фрагментов). Дополнительно ученые показали, что способность синтезировать ДНК на матрице РНК у Polθ (а также и у HIV RT) сохраняется при достаточно широком диапазоне условий (разных значениях pH и концентрации KCl). Полимераза Polη теряла такую способность при малейшем отклонении условий от оптимальных значений. Ни у одной другой ДНК-полимеразы человека (а проверили все восемь имеющихся у человека ДНК-полимераз) этой способности не выявлялось вовсе.
Ученые также показали и субстратную специфичность обратной транскриптазы ВИЧ — она могла эффективно работать только на матрице РНК, но с большим трудом строила фрагменты на матрице ДНК (длинные фрагменты получались только при очень высоких концентрациях фермента). А вот Polθ действительно эффективно работала и на матрице ДНК, и на матрице РНК.
Следующая серия экспериментов показала, что, во-первых, Polθ обладает более высоким сродством к гибридным молекулам ДНК/РНК, чем к ДНК/ДНК, во-вторых, синтез ДНК на матрице РНК идет и быстрее, и точнее, чем на матрице ДНК. Частично результаты этих экспериментов показаны на рис. 3.

Рис. 3. Скорость и точность синтеза полимеразой тета ДНК на матрицах РНК или ДНК. А — варианты субстратов, использованных в экспериментах — дуплексы, образованные ДНК-матрицей с ДНК-затравкой или РНК-матрицей с ДНК-затравкой (последовательности ДНК показаны черным шрифтом, последовательности РНК — красным). B — результаты серии экспериментов по оценке скорости включения нуклеотидов в ходе полимеризации ДНК. В ходе эксперимента в смесь помимо субстрата и фермента добавлялись меченные дезоксинуклеотиды только одного типа из четырех существующих (dCTP, dTTP, dATP или dGTP). Это был нуклеотид, комплементарный следующему за затравкой в матрице. Оценивали, какая доля затравок оказывалась помечена включением нуклеотида (% extension) в течение определенного промежутка времени (измеряемого в минутах, min). На всех графиках черные точки соответствуют экспериментам с ДНК-матрицей, а точки другого цвета — экспериментам с РНК-матрицей. Как можно заметить, скорость включения трех из четырех нуклеотидов была выше при работе на матрице РНК. C — результаты серии экспериментов по оценке точности включения нуклеотидов в ходе полимеризации ДНК. В реакционную смесь добавляли субстрат, фермент и меченные дезоксинуклеотиды единственного типа. Но теперь, наоборот, это был нуклеотид НЕ комплементарный следующему за затравкой в матрице: его включение обозначало ошибочное срабатывание фермента. Более высокой скорость удлинения затравки теперь оказалась для ДНК-матрицы, то есть фермент реже допускает ошибочное включение нуклеотида при работе на матрице РНК. Рисунок из обсуждаемой статьи в Science Advances
Вероятным объяснением различий в скорости и точности синтеза на ДНК- или РНК-матрицах является изменение конформации белка в зависимости от связываемого субстрата. Действительно, оказалось, что субдомены каталитического домена претерпевают достаточно заметные конформационные изменения при связывании дуплексов ДНК/РНК или ДНК/ДНК — это различие можно видеть на рис. 1. Оказалось, что несколько аминокислот в субдомене, именуемом «большой палец», образуют дополнительные водородные связи с рибозой при связывании гетеродуплекса ДНК/РНК, что, вероятно, и объясняет более высокое сродство фермента к субстрату такого типа, равно как и более высокую скорость и точность синтеза ДНК, чем при работе с субстратом типа ДНК/ДНК.
Все вышеописанные данные были получены при исследовании свойств белка вне живой системы — in vitro. Конечно, чтобы выводы были вполне убедительными, нужно было убедиться, что Polθ действительно может использовать РНК как матрицу, работая непосредственно в живой клетке в качестве починщика ДНК. Эксперименты оказались вполне успешными. Мы опишем лишь некоторые из них, выполненные на клетках человека.
Ученые работали с линией клеток остеосаркомы (опухоли кости) человека U2OS. В эти клетки был встроен репортерный ген GFP (кодирующий зеленый флуоресцентный белок). Работу такого гена можно видеть по свечению клеток. Однако в данном случае ген был «подпорчен» делецией семи нуклеотидов в середине последовательности, да еще и вставкой небольшого постороннего фрагмента сразу около делеции. Вторую тестовую линию — POLQe16m — получили на основе вышеописанной, добавив еще одну модификацию — в этой линии полностью вывели из строя ген Polθ.
Первый эксперимент состоял в том, что в клетки родительской линии U2OS с дефектным GFP вносили небольшую цепочку нуклеиновой кислоты, содержащий верную последовательность центральной части гена GFP — без инсерции и с возвращенными семью нуклеотидами. Этот фрагмент должен был служить матрицей для заделывания бреши в гене GFP. Для того, чтобы сделать брешь и запустить механизм репарации, использовали систему CRISPR/Cas, которую натравили на встроенный в ген GFP чужеродный участок.
Нуклеиновая кислота, которая должна была служить матрицей для починки хромосомы, представляла собой либо целиком одноцепочечную молекулу ДНК, либо содержала два рибонуклеотида, попадающие в те самые недостающие семь нуклеотидов внутри GFP. Свечение клеток в итоге могло появиться только в том случае, если при заполнении бреши будет верно «списана» последовательность соответствующей цепочки фрагмента для редактирования. Рис. 4 показывает, что свечение восстанавливалось в большинстве клеток при использовании как цепочки, полностью построенной из дезоксирибонуклеотидов, так и при использовании цепочки с вставкой из двух рибонуклеотидов. Если тот же эксперимент проделывали с линией POLQe16m, то восстановления свечения при введении цепочки с рибонуклеотидами не наблюдалось, а вот если в эту линию вносили плазмиду с геном Polθ, то возвращалась и способность чинить хромосому. Схема и результаты этих серий экспериментов показаны на рис. 4.
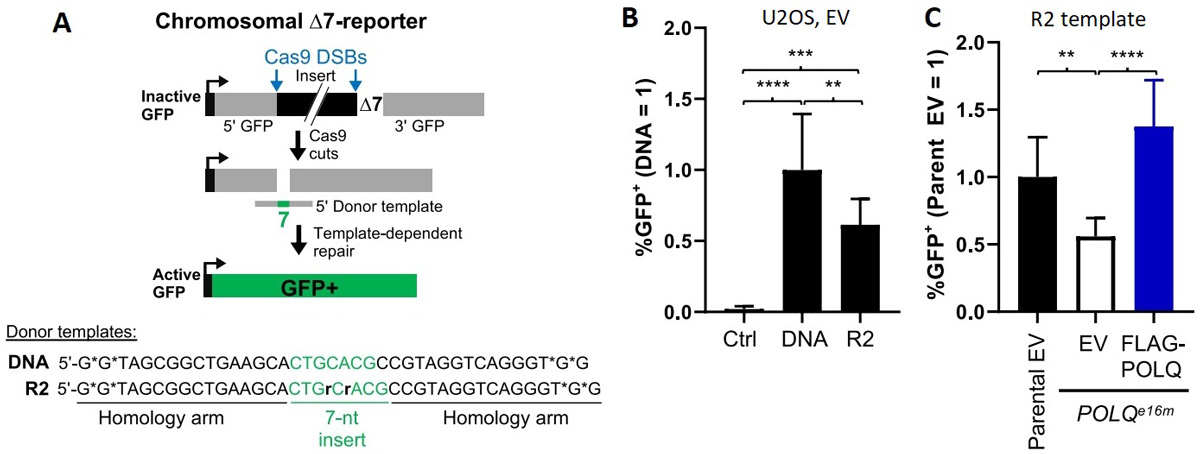
Рис. 4. Эксперименты на клетках человека. А — схема эксперимента. В клетки родительской линии U2OS или производной линии POLQe16m, каждая из которых несет в геноме дефектный ген зеленого флуоресцентного белка GFP, вносили компоненты системы CRISPR, нацеленной на GFP, чтобы внутри него образовался разрыв хромосомы. Одновременно вносили «донорную матрицу» (donor template), которая предположительно могла послужить матрицей при репарации разрыва, восстановив при этом ген GFP в его рабочем состоянии (active GFP). Донорная матрица представляла собой либо одноцепочечную ДНК (DNA), либо цепочку ДНК с включением по центру двух рибонуклеотидов (R2). В — результаты экспериментов на линии U2OS. По вертикальной оси показан процент клеток, которые флуоресцировали зеленым, — то есть в которых репарация прошла согласно задумке экспериментаторов. Ctrl — контрольные клетки, в которые вносили систему CRISPR, но не вносили донорную матрицу; следующие два столбика соответствуют клеткам с внесением разных вариантов донорных матриц. В обоих случаях в части клеток наблюдается восстановление функции гена GFP. При использовании ДНК-матрицы столбик получился выше — это неудивительно, поскольку репарацию с использованием ДНК матрицы могут осуществлять многие полимеразы в клетках человека, а репарацию с использованием смешанной матрицы — предположительно, только Polθ. С — сравнение результатов экспериментов на линях U2OS и POLQe16m при использовании донорной матрицы R2. Результат для линии U2OS здесь принят за 1 (первый столбик). Второй столбик показывает результат для линии POLQe16m (в ней отсутствует рабочий ген полимеразы тета), в которую вводили пустой вектор, а третий — для этой же линии, но с внесенным в составе искусственного вектора гена полимеразы тета (FLAG-POLQ). Разница между первым и вторым, а также между вторым и третьим столбиками доказывает, что именно полимераза тета ответственна за восстановление гена GFP при использовании донорной матрицы с рибонуклеотидами. Рисунок из обсуждаемой статьи в Science Advances
Присутствие в геномной ДНК участков, образованных гетеродуплексами ДНК/РНК было установлено достаточно давно. Это могут быть единичные встроенные рибонуклеотиды, а могут быть участки с достаточно большой протяженностью. Причины их образования, как и механизмы их устранения, бывают разными — этому вопросу посвящен большой недавний обзор G. M. Nava et al., 2020. One, No One, and One Hundred Thousand: The Many Forms of Ribonucleotides in DNA. Это может быть естественной частью некоторых молекулярно-биологических процессов (рис. 5): транскрипции, репарации, репликации (в частности, из рибонуклеотидов строятся фрагменты Оказаки — затравки для репликации запаздывающей цепи). Вместе с тем рибонуклеотиды достаточно часто ошибочно включаются ферментами в ходе синтеза ДНК при репликации или репарации (приблизительная частота таких ошибок 10−3), несмотря на высокую избирательность в пользу дезоксирибонуклеотидов. Это объясняется тем, что в наших клетках концентрация рибонуклеотидов всегда намного (до 100 раз) выше концентрации дезоксирибонуклеотидов. Впрочем, в клетках есть несколько видов охранных систем, которые настроены на выявление и удаление рибонуклеотидов из ДНК — и это очень важно для поддержания нормальной работы генов, репликации ДНК и сохранения ее в целости и сохранности.

Рис 5. Разнообразные механизмы появления дуплексов типа ДНК/РНК в хромосомном материале клеток. В клетках эукариот имеется обширный арсенал средств для устранения таких временно образующихся гетеродуплексов и восстановления нормальной структуры хромосом в форме двухцепочечной ДНК. Рисунок из обзорной статьи G. M. Nava et al., 2020. One, No One, and One Hundred Thousand: The Many Forms of Ribonucleotides in DNA
Но вот Polθ, исходя из результатов обсуждаемого исследования, обладает способностью использовать непосредственно рибонуклеотиды в качестве матрицы для синтеза ДНК. И эта ее способность пригождается на этапе репарации повреждений ДНК, по-видимому, главным образом тогда, когда по тем или иным причинам забуксовали другие охранные механизмы.
Таким образом, у млекопитающих впервые обнаружили фермент репарации с несколькими уникальными свойствами, включающими особенности структурной пластичности, способность прочно связывать гетеродуплексы ДНК/РНК и способность быстро и сравнительно с высокой точностью осуществлять обратную транскрипцию, выстраивая ДНК на матрице РНК. Это объясняет, каким образом наши клетки справляются с одним из достаточно частых нарушений в структуре ДНК, когда часть цепочки оказывается построена не из дезоксирибонуклеотидов, а из рибонуклеотидов.
Источник: Gurushankar Chandramouly, Jiemin Zhao, Shane McDevitt, Timur Rusanov, Trung Hoang, Nikita Borisonnik, Taylor Treddinick, Felicia Wednesday Lopezcolorado, Tatiana Kent, Labiba A. Siddique, Joseph Mallon, Jacklyn Huhn, Zainab Shoda, Ekaterina Kashkina, Alessandra Brambati, Jeremy M. Stark, Xiaojiang S. Chen and Richard T. Pomerantz. Polθ reverse transcribes RNA and promotes RNA-templated DNA repair // Science Advances. 2021. DOI: 10.1126/sciadv.abf1771.
Татьяна Романовская


А так же там говорилось, что после вакцинации люди стали притягивать металлические предметы, разъяснялся тайный смысл речей Трампа и предлагалось купить амулетов, защищающих от 5Г, книг о заговоре рептилоидов и постеров с лицом Била Гейтса.
Забавно, что всё это изобрели в Англии, стране, где возникла наука.
-
Однако откуда все эти побочные эффекты в том числе смертельные не способны объяснить создатели вакцин, разные светилы, международные и местные чиновники, и предсказать это все тоже ниасилили. Да и вообще зачем объяснять, не каждый же умирает, все в порядке - зелье безопасно. Надо ли использовать и как тоже мнения часто меняются, да еще деньги думать мешают - руки жгут, дурманят шелестом, и политическая игра идет без остановки - надо с козыря ходить.
-
-
Убивает 1%, в самом деле ковидла очень избирательная, при том что "лечить" вакцинами предлагается, или заставляют, всех здоровых людей, в том числе тех кого вирус не способен поразить, как по неизвестным причинам, так и по известным - была информация что курение табака защищает, но это абсолютно игнорируется, как и очевидная необходимость наглухо закрыть границы и чтобы нового привозного вируса точно не было.
-
Так умирают или тяжело болеет не только больные. Как гены совпали. А кто то с лишним весом и возрасте, а коронавирус не берёт.
Пару смертей на сотни миллионов европейцев вполне допустимая цифра, учитывая эффект от коронавируса. Да вакцины от полиомиелита то же не долго проверяли. На это были причины.
Другое дело, что даже если эпидемия кончится, тоталитарные меры по контролю власти будут отменять очень неохотно, а то и оставят на совсем. И после заточение ещё долго население не сможет отойти.
-
-
-
Особливо - полиадениновая вставка тудыть...
Также интересно в этой связи проверить "судьбу" кучи РНКовых продуктов (например, в семенной жидкости)...
-
Бил Гейтс, наверное, об этом уже знал. Да ещё телегония может работать. Подозреваю, скоро узнаем, что энзим активизируется при электромагнитном излучении, используемого в 5Г. И прочая народная мудрость...
И в итоге человечество разделится на две части, те, которые имеют ген шпиля капсида, и те, которые не имеют. Так скажем пацаки и чатлане, мрнк+ и мрнк-. И у всех на лбу будет QR код.-
Вы так активно пытаетесь остроумничать, что прям тошнотно становится. Просто не только лишь не все имеют хорошее чувство юмора, но мало кто имеет... И вы не из их числа, увы и ах. Это точно. :]
энзим активизируется при электромагнитном излучении
Во-первых, футакговорить! А во-вторых, таки это возможно, можете начинать просвещаться отсюдова (и далее везде): http://dx.doi.org/10.1021/acsnano.7b06439-
таки это возможно, можете начинать просвещаться отсюдова (и далее везде): http://dx.doi.org/10.1021/acsnano.7b06439
Коментаторы в ступоре.
Но не следует паниковать раньше времени.
В статье описывается магнитомеханическое разрушение комплекса ингибитора с ферментом при низких частотах.
Как хорошо, что это не модифицированный криптохром переключающийся на определенной частоте СВЧ излучения.-
Коментаторы в ступоре.
А я и не говорил, что будет легко... :Dмагнитомеханическое разрушение комплекса
Два чая этому господину! :)на определенной частоте СВЧ излучения
У хитропопых моделистов есть такая штука: "Steered molecular dynamics"... Наподобие такого: https://doi.org/10.1038/s41598-020-70863-9
Нужно всего лишь связать типовые частоты бытовых СВЧ излучателей с (прикладываемыми) тянитолкайными силами, и вуаля! :з-
Молекулярная динамика вторична. Здесь рулит квантовая механика.
Слабые магнитные поля влияют на живых существ.
https://biomolecula.ru/articles/nobel-vs-shnobel-ili-mekhanizmy-magnitoretseptsii
Но это не критично.
Однако нашлась в инете статья, подтверждающая мои худшие опасения - управление по Wi-Fi, то есть без вживления чипов и электродов:
http://csr.spbu.ru/pub/RFBR_publications/articles/chemistry/2001/spinovaya_himiya_01_chem.pdf Микроволновый МИЭ — это также красивый способ контролировать химическую реакционную способность, управлять ею с помощью селективной микроволновой манипуляции электронным спином реагентов.
Немагнитное происхождение спинового катализа позволяет ему эффективно влиять на химическую динамику пар радикалов даже в отсутствие магнитных полей и сверхтонких электрон-ядерных взаимодействий.
Ясно, что ионы и комплексы парамагнитных металлов, будучи химическими катализаторами, одновременно могут выполнять роль спиновых катализаторов, разрешая протекание спин-запрещенных процессов.
Практическое использование спиновой химии — дело ближайшего будущего в наступившем веке.-
Как быстро спин релаксирует в этих металло-комплексах? Поди, постоянно надо подзаряжать клиента, чтоб метода работала.
-
Как быстро спин релаксирует в этих металло-комплексах? Поди, постоянно надо подзаряжать клиента, чтоб метода работала.
Судя по отсутствию нобелей за прошедшие двадцать+ лет, метода не заработала совсем. Ну или власти скрывают от активной апчественности, как обычно... :Р
Ящитаю, ВайФаем надо жахать сразу по лептонам - вот где золотая жила! :]-
Ящитаю, ВайФаем надо жахать сразу по лептонам - вот где золотая жила! :]
Ученых учить - только портить.
Жахают родимые, жахают уже давно.
https://pereplet.ru/obrazovanie/stsoros/465.html
Куда в следущий раз будет введен ЭПР зонд, и какой каскад реакций будет запущен при передаче сигнала 5-G ?Спиновые метки - химически стабильные парамагнитные молекулы, которые используются в качестве молекулярных зондов для изучения структуры и молекулярной подвижности различных физико-химических и биологических систем.
Спиновую метку "загружают" внутрь клеток,...
На рис. 6, Б показан спектр ЭПР спиновой метки ТЕМПОН в клетках зародыша семян пшеницы.-
Ученых учить - только портить.
Ну вот так всегда!.. Только придумаешь что-то достойное (как подсократить число ртов..), а оно уже давно придумано... :/
Жахают родимые, жахают уже давно.Куда в следущий раз будет введен ЭПР зонд, и какой каскад реакций будет запущен при передаче сигнала 5-G ?
В моск, конечно же, - тут и думать не надо! :З
Причём сделать так, шоб поциэнты сами валом валили... Ну и назвать как-нибудь звучно.. например, фМРТ... :]-
Ну вот так всегда!.. Только придумаешь что-то достойное (как подсократить число ртов..), а оно уже давно придумано... :/
Все придумано до нас. В том числе антинаучный бред, прикрывающий передовые разработки.В моск, конечно же, - тут и думать не надо! :З
Мозг большой. Скорее всего в синапсы управляющих центров - например в гиппокамп.
Это будет песня:
https://youtu.be/pX06wlCAcUI?t=125Причём сделать так, шоб поциэнты сами валом валили... Ну и назвать как-нибудь звучно.. например, фМРТ... :]
Поскольку технология еще не отработана, повторять процедуру каждый год, а лучше каждые 6 месяцев.
-
-
-
-
-
Как быстро спин релаксирует в этих металло-комплексах? Поди, постоянно надо подзаряжать клиента, чтоб метода работала.
Естественно. Это ведь не детекторный приемник.
СВЧ антеной могут быть радикалы или триплет возбужденные молекулы.
https://duckduckgo.com/?q=долгоживущие+радикалы&ia=web
https://xumuk.ru/encyklopedia/2/3786.htmlРадикал
https://ru.wikipedia.org/wiki/Флуоресценцияы свободные могут быть короткоживущими (время жизни доли секунды) или долгоживущими (до неск. лет), нейтральными или заряженными (см. Ион-радикалы), иметь один или большее число неспаренцых электронов (напр., бирадикалы), быть твердыми, жидкими или газообразными в-вами.
Осн. специфич. методы обнаружения и исследования строения радикалов свободных основаны на использовании спектроскопии электронного парамагнитного резонанса.
Синглет-триплетные переходы имеют квантовомеханический запрет, поэтому время жизни возбуждённого состояния при фосфоресценции составляет порядка 10−3 − 10−2 с.
-
-
-
-
-
-
-
2) еще и очень сильно у них меняется скорость протекания в зависимости от изотопного состава идущих в комплексе ионов магния, точнее как они посчитали ядерного спина
Кинетическому изотопному эффекту уже лет сорок-писят. :)
Токма промерять его могли/могут на субстратах/продуктах (ибо они выступают в роли "челноков"). :D
-
1я часть - ин витро. здесь остаётся только удивляться, как такой эффект оставался незамеченным столько лет, ибо точность работы этой полимеразы в ОТ-режиме намного выше, чем в ДНК-зависимом (см рис 3). это случайность, или отбор продавил такую особенность?
2я часть - ин виво, вообще про другое. там полимераза справилась с наличием двух небольших химических изменений в матрице - по одному кислороду на мономер. здесь мало новизны, и так было известно, что она мало чувствительна к подобным "ошибкам". возникает другой вопрос - может ли она в клетке по мРНКе синтезировать ДНКу? и как к этому отнесутся рестриктазы, не воспримут ли они это как вирусную атаку?
Новости: Молекулярная биология






Рис. 1. Модели пространственной структуры комплекса из полимеразы тета (Polθ) с дуплексом ДНК/ДНК + дидезоксиГТФ (слева) и с дуплексом ДНК/РНК + дидезоксиГТФ (справа). Непосредственный участок взаимодействия белка с присоединяемым нуклеотидом дидезоксиГТФ (ddGTP) показан дополнительно во врезках. В каталитическом домене полимеразы выделяют субдомены «ладонь» (palm), «пальцы» (fingers) и «большой палец» (thumb). Рисунок из дополнительных материалов к обсуждаемой статье в Science Advances