Определена структура малой субъединицы эукариотической рибосомы

Рибосомы — это большие молекулярные машины, состоящие из молекул РНК и белков, которые присутствуют во всех живых клетках. Их биологическая роль заключается в осуществлении трансляции — процесса биосинтеза белковых молекул, в ходе которого последовательность аминокислот в полипептидных цепях однозначно определяется информацией генетического кода. В 2000 году ученые определили атомную структуру малой субъединицы бактериальной рибосомы и большой субъединицы рибосомы архебактерий. Это позволило гораздо лучше понять работу трансляционного аппарата у прокариотических организмов. Теперь удалось определить молекулярную структуру малой субъединицы эукариотической рибосомы в комплексе с фактором инициации трансляции eIF1 с разрешением 3,9 ангстрем.
Рибосомы всех живых организмов состоят из двух субъединиц — малой субъединицы, которая декодирует информацию с молекул матричных РНК (мРНК), и большой субъединицы, которая катализирует образование пептидной связи между растущей полипептидной цепочкой будущего белка и каждой последующей аминокислотой. По сравнению с рибосомами бактерий эукариотические рибосомы гораздо крупнее (около 4300 кДа) и сложнее в организации (имеют дополнительные 45 белков и две рибосомальные РНК). Хотя их основные функции довольно консервативны у всех живых организмов, тем не менее процесс инициации белкового синтеза у эукариот существенно отличается от аналогичного процесса прокариот.
Стадия инициации белкового синтеза у эукариот является регуляторной мишенью в ряде клеточных процессов, включая развитие, процессы дифференцировки, реакцию на стресс и нейронное функционирование (то есть на этой стадии происходит многосторонняя регуляция этих процессов). Многие заболевания, включая рак и нарушения метаболизма, связаны с нарушением регуляции инициации трансляции. Центральную роль в этом процессе играет малая рибосомальная субъединица (40S) — она связывается с факторами инициации, которые запускают сканирование мРНК и начало белкового синтеза. Причем если в бактериальном аппарате трансляции используется только три фактора инициации, то трансляция эукариот зависит как минимум от 12 факторов инициации, которые упорядоченно собраны в малой субъединице. Также необходимо отметить, что, в отличие от бактерий, у эукариотических организмов сборка рибосом происходит в разных клеточных компартментах.
Группе ученых из Института молекулярной биологии и биофизики (Цюрих, Швейцария) удалось кристаллизовать комплекс малой 40S-субъединицы эукариотической рибосомы и фактора инициации eIF1 и определить кристаллическую структуру с разрешением 3,9 ангстрем.
Как известно, биомолекулы способны образовывать кристаллы, которые могут рассеивать рентгеновские лучи. На основании картины дифракции рентгеновских лучей кристаллами можно воссоздать трехмерную кристаллическую структуру. Процесс получения трехмерной структуры белка состоит из нескольких стадий. Поскольку для кристаллизации необходимо большое количество белка, то, как правило, первым этапом является молекулярное клонирование его гена (генов) с последующей экспрессией, например в специальных бактериальных штаммах. Далее происходит очистка белкового препарата в несколько стадий различных хроматографий, так как белки, используемые для кристаллизации, должны обладать высокой степенью чистоты. Следующая стадия — кристаллизация белков (или комплексов), где применяется довольно много технических ухищрений для получения кристаллов, подходящих для дальнейшей работы. После получения удачных образцов кристаллов их замораживают и хранят в жидком азоте до следующей стадии — рассеивания рентгеновских лучей и получения картины их дифракции. Этот этап осуществляется при помощи синхротронов: кристалл облучается рентгеновскими лучами при разных углах поворота вокруг собственной оси. Картины дифракции записывают и анализируют при помощи специализированного программного обеспечения. Одна из главных характеристик на этом этапе — разрешение дифракционной картины рассеивания рентгеновских лучей, что определяет, например, будут ли видны отдельные аминокислоты на конечной структуре или только элементы вторичной структуры белка. Последняя стадия — построение компьютером модели структуры белка и окончательное определение пространственной структуры белка или комплекса белков с другими молекулами.
Благодаря комбинации методик по моделированию атомных структур, авторы построили пространственную модель рибосомальной 40S-субъединицы, а также определили третичную структуру всей молекулы 18S-рибосомальной РНК (рРНК); см. рис. 1.

Эукариотическая 40S-субъединица содержит 33 белковые молекулы, 18 из которых отсутствуют у бактерий. В результате данного исследования были показаны пространственные укладки всех рибосомальных белков малой 40S-субъединицы (рис. 2 и 3).
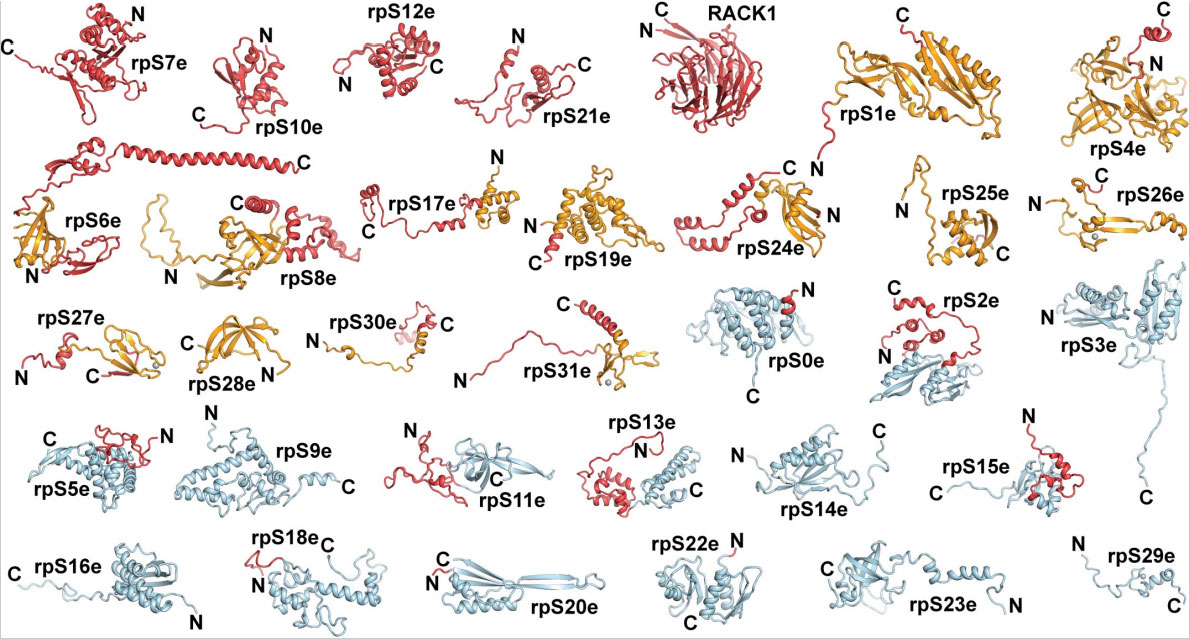
К настоящему моменту известно, что для преодоления пространственных помех во время процесса трансляции, обусловленных элементами вторичной структуры мРНК, рибосомы обладают внутренней хеликазной активностью, чтобы расплетать молекулы мРНК. В ходе исследования был смоделирован канал для мРНК в эукариотической рибосоме, особенности которого по сравнению с аналогичной структурой бактериальных рибосом также обуславливают разницу инициации трансляции у прокариот и эукариот (рис. 4A).
Теперь известно, что эукариотические рибосомальные белки образуют протяженные контакты между собой, а не с рРНК, как у прокариот. Полученные данные позволяют говорить о том, что форма «клюва» эукариотической малой субъединицы похожа на такую же структуру у малой субъединицы прокариотической рибосомы, но состоит в основном из белков, а не из РНК (рис. 4B).

B) Модель структурного элемента «клюв» эукариотической рибосомы. У эукариот молекула рРНК в структурном элементе «клюв» редуцирована и содержит мотив «изгиб-поворот», к которому присоединен белок rpS12e. Ниже показан структурный элемент «клюв» бактериальной рибосомы, который преимущественно сформирован молекулой рРНК. Иллюстрация из обсуждаемой статьи в Science
Эукариотический фактор инициации eIF1 не имеет структурных гомологов у бактерий, хотя некоторые функции у него схожи с бактериальным фактором инициации трансляции IF3. Фактор eIF1 играет важную роль в инициации трансляции у эукариот — с его участием происходит переход 40S-субъединицы в конформацию, готовую к сканированию мРНК. Во время сканирования фактор eIF1 способен распознать стартовый кодон трансляции, далее происходит его диссоциация из инициаторного комплекса, что приводит к формированию закрытого комплекса у малой 40S-субъединицы. В данной работе авторы впервые показали точную локализацию и характер взаимодействия инициаторного фактора eIF1 с 40S-субъединицей (рис. 5), что приоткрывает занавес над вопросом о взаимодействии фактора eIF1 и инициаторной транспортной РНК (тРНК).

Таким образом, определение кристаллической структуры 40S-субъединицы демонстрирует функциональные и эволюционные особенности эукариотических рибосом. Для дальнейшего уточнения полученных данных и ответов на вопросы о сборке, созревании и транспортировке 40S-субъединиц необходимы биохимические и генетические эксперименты, отправной точкой для которых послужит полученная в результате данной работы структура.
Источник: Julius Rabl, Marc Leibundgut, Sandro F. Ataide, Andrea Haag, Nenad Ban. Crystal Structure of the Eukaryotic 40S Ribosomal Subunit in Complex with Initiation Factor 1 // Science. 11 February 2011. V. 331. P. 730–736.
См. также:
Тайна происхождения рибосом разгадана?, «Элементы», 27.02.2009.
Максим Нагорных


Новости: Молекулярная биология






