Вековая история одной палеоботанической идеи
Сергей Наугольных
«Природа» №7, 2020
 Об автореСергей Владимирович Наугольных — доктор геолого-минералогических наук, главный научный сотрудник Геологического института РАН, профессор РАН; контрактная аффилиация — ведущий научный сотрудник научно-исследовательской лаборатории стратиграфии Казанского федерального университета. Область научных интересов — палеоэкология, систематика и эволюция высших растений, изучение палеопочв. |
В науках о прошлом Земли время от времени складываются мифологемы, которые благодаря своей кажущейся логичности и стройности становятся очень популярными и широко тиражируются, в особенности журналистами и средствами массовой информации. Когда же появляются новые данные, меняющие или даже опровергающие такую мифологему, изменить ее чрезвычайно трудно. Такова и вековая история о «русских глоссоптерисах» профессора Владимира Прохоровича Амалицкого. Но с чего же в действительности началась эта история?
Магнетизм предконцепции
Вот цитата из одной книги моего палеонтологического детства, ее я частенько листаю просто для удовольствия, и стоит она на «золотой» полке моей библиотеки: С неослабевающей энергией ученый продолжал поиски в последующие годы. Новая ценная находка обогатила его коллекции: в пермских слоях Амалицкий обнаружил хорошо сохранившиеся отпечатки листьев древних семенных растений — папоротников глоссоптерисов. Эти мощные древовидные растения также росли в пермское время в Южной Африке. Судя по отпечаткам, глоссоптерисы, жившие на севере, ничем не отличались от южноафриканских1 [подчеркнуто мной. — С.Н.].
Именно так в сжатом виде и звучит миф о глоссоптерисах, найденных Амалицким в пермских отложениях на р. Северной Двине. Давайте попытаемся разобраться во всех деталях этой совсем не простой палеоботанической истории более чем вековой давности. Но сразу же сделаю оговорку: глоссоптерис — растение голосеменное, и никакого отношения к папоротникам он не имеет, вопреки не совсем грамотным сведениям в некоторых научно-популярных книгах.
В чем главная цель, или даже, если уместно так выразиться, идея этой статьи? Хочу в силу своих способностей показать прихотливость путей познания далекого прошлого Земли, и неоднозначность выводов, к которым может прийти исследователь, опираясь на внешнее сходство своих находок с широко известными окаменелостями. Конечно, нельзя винить предшественников в том, что результаты их исследований сейчас приходится пересматривать. Но и на современных ученых специфика палеонтологического знания накладывает большую ответственность, требующую глубокого анализа и своих собственных данных, и результатов, полученных коллегами. Как увидит читатель, точка в истории изучения российских глоссоптерисов пока не поставлена, но и то, что мы знаем сейчас, позволяет по-новому взглянуть на историю позднепалеозойского растительного мира.
Клише «первое упоминание о «русских глоссоптерисах» принадлежит именно Амалицкому» широко распространено среди любителей палеонтологии. На самом деле впервые о находках листьев глоссоптерис в России за полвека до Амалицкого — в далеком 1840 г. — написал один из первых российских палеонтологов Г. И. Фишер фон Вальдгейм, основатель Московского общества испытателей природы. В своей работе Фишер фон Вальдгейм упомянул о находке листьев двух видов глоссоптериса: Glossopteris phillipsii и G. crenulata (второй вид был определен со знаком вопроса) в верхнепермских отложениях Оренбуржья, на Южном Урале [1]. Спустя десятилетие о находках G. crenulata написал выдающийся российский естествоиспытатель того времени К. Мерклин [2], специалист по ископаемым древесинам, работы которого получили мировую известность.
Таким образом, присутствие ископаемых листьев «глоссоптерисового» облика в пермских отложениях европейской части России и в Приуралье отмечалось отечественными учеными задолго до начала первых экспедиций Амалицкого. Надо полагать, что данные, опубликованные Фишером фон Вальдгеймом, вполне могли лечь в основу представлений Амалицкого о связях между Гондваной и Ангаридой, к западным пределам которой в пермском периоде относилась и нынешняя территория Европейской России. О возможности такой связи, кстати сказать, писал гениальный швейцарский геолог Э. Зюсс в фундаментальной трехтомной работе «Лик Земли» (1883–1888). У меня нет особых сомнений в том, что, работая в Санкт-Петербурге и Варшаве, Амалицкий не мог не ознакомиться с «Ликом Земли» и, более того, наверняка использовал его в своей преподавательской деятельности.
Зюсс в «Лике Земли» [3, p. 389] специально останавливается на характеристике пермской биоты Южной Африки (центральной части Гондваны). Говоря о фауне позвоночных, попутно он вспоминал и о растениях, называя находки в песчаниках верхней части формации Кару (слои Бофорт) листьев Glossopteris browniana побегов хвощевидных Phyllotheca indica. Листья и побеги филлотек отмечались и для каменноугольных и пермских отложений Ангариды, но эта тема, пожалуй, заслуживает отдельной статьи. В той же части книги Зюсса, помимо упоминания южноафриканских глоссоптерид, присутствует указание на возможную синхронность пермских тиллитов (ледниковых отложений) Северо-Западной Европы (Англии) и Южной Африки.
Но не только от геологов поступали сведения о возможных связях северного континента Ангариды (Зюсс называл его континентом Ангара) с материками Гондваны. Так, в 1879 г. один из первых российских палеоботаников И. Ф. Шмальгаузен описал из нижнепермских отложений (по современным представлениям, из бургуклинской свиты) Тунгусского бассейна голосеменное растение, которое назвал Zamiopteris glossopteroides. Шмальгаузен не случайно дал новому растению видовой эпитет glossopteroides, т.е. имеющий сходство с глоссоптерисом. Действительно, листья нового растения имели языковидные или ланцетовидные очертания, оттянутое клиновидное основание, вееровидное жилкование с ложной средней жилкой или осевым пучком жилок, расположенным в средней (медиальной) части листа. Внешне эти листья действительно напоминали листья глоссоптерид. Здесь уместно отметить, что спустя шестьдесят лет после выхода в свет работы Шмальгаузена, из пеляткинской свиты Тунгусского бассейна были описаны листья Pursongia tunguscana [4], которые достоверно обладали анастомозами и, таким образом, вполне соответствовали диагнозу рода Glossopteris.
Продолжая отстаивать сходство позднепалеозойской ангарской флоры с флорой Гондваны, другой выдающийся российский палеоботаник М. Д. Залесский писал: Suess исходил из той мысли, что развитие органической жизни на континенте Гондвана на юге и на континенте Ангара на севере, несмотря на присутствие между ними моря, могло следовать параллельно на обоих материках, так как для него уже было ясно некоторое отношение флоры Ангары к флоре Гондванского континента [5, с. 5]. И далее: Настоящая работа является попыткой объединить в одно целое все элементы флоры, найденные на громадной площади Северной Азии из палеозойских осадков Зюссовского континента Ангара, элементы, по которым эта флора нам кажется родственной или сходной по характеру своему с гондванской флорой Индии и других частей гондванской суши и является, по-видимому, по возрасту пермской. В настоящее тревожное время, переживаемое этой страной, я принужден опубликовать только атлас этой интересной флоры в сопровождении объяснений к таблицам его, но, когда в стране представится возможность более спокойной работы, за ним последует текст, в котором будет представлено как описание изображенной флоры, так и критическое ее изучение, а затем будут приведены те геологические выводы, к которым можно прийти из сравнительного изучения ее со сходными флорами [5, с. 6]. К вопросу о связах ангарской и гондванской флор Залесский обращался неоднократно [6–9].
В 1918 г. Залесский, конечно же, не догадывался, что Россию ждут еще более суровые испытания и что сам он будет интернирован 24 июля 1943 г. из Орла, где он тогда жил и трудился, в фашистскую Германию, в Берлин, в Имперский институт изучения почв, и что домой он сможет вернуться только после окончания войны2.

Листья Zamiopteris glossopteroides из нижнепермских отложений Тунгусского бассейна [23, табл. XIV, фиг. 1]
Показательно, что в работе 1918 г. Залесский перенес вид Zamiopteris glossopteroides непосредственно в состав гондванского рода Gangamopteris, считая, что листья из Тунгусского бассейна обладают анастомозами, характерными для глоссоптерид [5, с. 24].
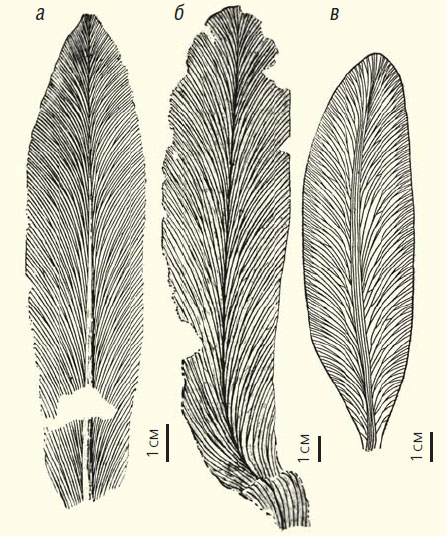
Однако по мере изучения пермских флор Ангариды морфологическое своеобразие и особенности строения ангарских растений все больше бросались в глаза Залесскому, поэтому часть видов из ранее определявшихся гондванских таксонов он отнес к новым родам. Так, для ланцетовидных листьев из верхнепермских отложений севера Европейской платформы и сходных с ними листьев Приуралья и Сибири Залесский предложил новый род Pursongia. Пурсонгия, тем не менее, рассматривалась Залесским в составе порядка Glossopteridales.
Новые взгляды
К проблеме родственных связей и таксономического положения рода Zamiopteris и морфологически близких к нему растительных остатков в середине 60-х годов прошлого века обратился С. В. Мейен, известный палеоботаник и эволюционист. Мейен предложил дополненный и исправленный диагноз рода Zamiopteris [10, с. 86], в котором подчеркнул отсутствие анастомозов между жилками и конспективно привел эпидермально-кутикулярные признаки рода. В указанной статье он справедливо отметил, что нет никакой гарантии, что эпидермально-кутикулярное строение типового вида Zamiopteris glossopteroides из перми Тунгусского бассейна и описанного им нового вида Z. neuburgiana было однотипным, поэтому нет уверенности, что эти два вида действительно принадлежали к одному и тому же роду. Правда, при этом эпидермально-кутикулярные признаки Z. neuburgiana почему-то с легкостью были перенесены в расширенную диагностику рода Zamiopteris. Таким образом, в словах Мейена и принятом им таксономическом решении наблюдается очевидное противоречие.
Мейен сближал род Zamiopteris с кордаитами, однако из описания как макроморфологических, так и эпидермально-кутикулярных признаков вида Z. neuburgiana это никак не следует. На мой взгляд, Zamiopteris — это голосеменное растение, родственное пурсонгиям, но более древнее и менее эволюционно продвинутое. Фрагментарные листья, описанные Мейеном как Zamiopteris sp. 1 и Zamiopteris sp. 2, по моему мнению, не могут быть отнесены к этому роду. Скорее это плохо сохранившиеся кордаитоподобные листья рода Rufloria.

Pursongia amalitzkii, один из экземпляров, найденных В. П. Амалицким в местонахождении Соколки. Образец из экспозиции Палеонтологического музея имени Ю. А. Орлова РАН, Москва
В той же работе Мейен подробно остановился и на статусе рода Pursongia. Вот цитата из его работы: Сразу оговоримся, что типовой материал Pursongia не сохранился, нет и его фотоизображений. Поэтому составить представление об облике листьев, относимых Залесским к Pursongia, можно только по образцам (с этикетками Залесского), имеющимся в Центральном геологическом музее имени Ф. Н. Чернышева в Ленинграде и в Палеонтологическом музее имени Ю. А. Орлова в Москве [10, с. 93].
Мейен доказал, что пурсонгиеподобные листья из верхней перми Русской платформы и Приуралья, которые он относил к особому роду Tatarina, ассоциативно связаны с репродуктивными органами Peltaspermum в форме семеносных дисков — пельтоидов, что доказывало принадлежность этих растений к семейству Peltaspermaceae порядка Peltaspermales, который сейчас нередко обособляют в отдельный класс Peltaspermopsida.
В отношении номенклатуры пурсонгиеподобных листьев необходимо отметить, что Мейен и его последователи попали в очень коварную логическую ловушку. Если мы признаем, что каждая специфическая форма сохранности или каждая новая возможность наблюдать какую-то особую группу признаков, недоступную для наблюдения на типовом экземпляре, — достаточное основание для введения в палеоботаническую номенклатуру новых таксонов (в данном случае — на уровне рода), то тогда мы открываем самый настоящий номенклатурный ящик Пандоры. К такой ситуации вполне подходит еще один метафорический образ — выпускание злобного джинна из бутылки, ведь в таком случае каждый желающий самоутвердиться на почве таксонотворчества получает формальные и легальные основания для неконтролируемой деятельности. Именно поэтому, когда в мое распоряжение попали ланцетовидные листья из верхнепермских отложений, обнажающихся у г. Вязники (Владимирская обл.), с жилкованием, характерным для пурсонгий, и с хорошо сохранившимися кутикулами [11], я описал их в качестве нового вида рода Pursongia, отдавая законный приоритет в изучении этого рода Залесскому. Заслуги Мейена в изучении ланцетовидных листьев из верхней перми Русской платформы и Приуралья отражены в видовом названии нового растения — Pursongia meyenii.
История изучения рода Pursongia со всеми перипетиями, которые ее сопровождали, проанализирована мной в отдельной статье, вошедшей в юбилейный золотой (пятидесятый!) том международного журнала Palaeobotanist [12], поэтому здесь ее повторять не буду. Но основные выводы необходимо кратко обсудить.
Какими были глоссоптериды?
Роду Glossopteris и его ближайшим родственникам посвящен гигантский массив литературы, так что привести даже краткий ее обзор в научно-популярной статье совершенно невозможно. Желающих окунуться в тему можно переадресовать к моему очерку, опубликованному несколько лет назад на страницах «Природы»3.
Собственно говоря, род глоссоптерис был первоначально предложен только для ланцетовидных листьев с характерным сетчатым жилкованием, образованным жилками, вееровидно расходящимися от осевого пучка жилок или от так называемой ложной средней жилки, и с анастомозами, соединяющими соседние жилки. Было описано огромное количество видов глоссоптерисов с самыми разными вариантами жилкования, в основе своей все же повторяющих базовую схему. Жилки могли быть и тонкими, и потолще, расходящимися к краям листа под разными углами. Анастомозы могли располагаться чаще или реже. И даже форма листа могла меняться от лентовидной до широкоязыковидной. Однако общий облик листьев, как правило, сразу давал понять исследователю, ископаемые остатки какого именно растения он нашел, неважно, случилось ли это в Индии, Африке, Южной Америке, Австралии или даже Антарктиде.

Реконструкция расположения семеносного органа Partha sp. на листе Glossopteris sp.: а — комбинированная фотография листа, б — реконструкция, рисунок автора
Репродуктивные органы глоссоптерисов (и глоссоптерид в целом) тоже имели более или менее общий план строения. И семеносные (женские), и производящие пыльцу (мужские) органы глоссоптерид прикреплялись к осевой части листа, образуя так называемые фертилигеры. Женские фертилигеры могли продуцировать либо большое количество мелких семян, падавших непосредственно рядом с материнским растением, либо несколько крупных семян, снабженных крылаткой, которые могли переноситься ветром на большее расстояние от места исходного произрастания. По моему мнению, это было связано с различной репродуктивной стратегией глоссоптерид. Часть из них была адаптирована к широкому расселению в условиях низкой конкуренции за ресурсы, а другая — к максимально большому производству диаспор в условиях плотного заполнения экологических ниш в растительном сообществе. Этот, по своей сути поведенческий, этологический феномен получил название «супраидиоадаптация» [13]. По форме роста среди глоссоптерид были и кустарники, и высокие деревья. Многие глоссоптериды, по всей видимости, стали важными углеобразователями на материках Гондваны.

Строение женских репродуктивных органов Peltaspermum polyspermis, принадлежавших пельтаспермовому с листьями Pursongia sp. из местонахождения Завражье: а — два соседних семеносных диска, один из которых виден полностью, на радиальных секторах хорошо сохранились овальные семенные рубцы; б — фрагмент кистевидного собрания семеносных дисков; в — продольное сечение через ножку и основание семеносного диска, сохранившихся в форме отпечатков
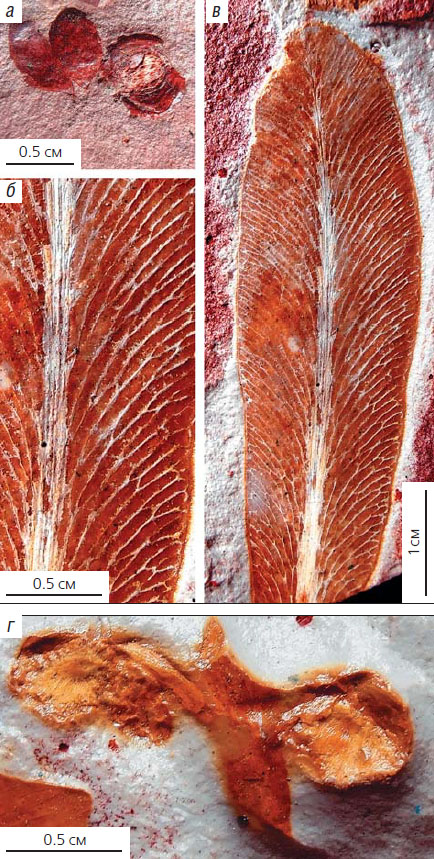
Глоссоптериды из верхнепермских отложений Австралии (угольное месторождение Иллавара, бассейн Данеду, Новый Южный Уэльс): а — раскрытый семеносный орган; б, в — листья Glossopteris sp.: б — сетчатое жилкование с анастомозами, в — общий вид листа, г — семеносный орган Partha sp.
Среди растений, отнесенных Залесским и другими палеоботаниками к роду Pursongia, оказались как листья без анастомозов (такие, как типовой вид рода P. amalitzkii), только внешне сходные с глоссоптеридами, так и листья с анастомозами (например, P. asiatica или P. tunguscana). Решить проблему установления точного таксономического положения ангарских пурсонгий с анастомозами, не располагая данными о репродуктивных органах, пока не представляется возможным.
Если кратко резюмировать значение различий пурсонгий и настоящих глоссоптерисов, то прежде всего следует отметить два момента. Во-первых, пурсонгии не имеют никакого отношения к глоссоптеридам, они принадлежат к совершенно иной группе голосеменных (а именно к пельтаспермовым) и не могут служить свидетельством в пользу палеофитогеографических связей между Гондваной и Ангаридой, как это предполагалось вначале. Во-вторых, при этом присутствие настоящих Glossopteris в пермских отложениях Сибири, Дальнего Востока и Монголии доказывает, что миграционные обмены между Гондваной и Ангаридой в течение пермского периода все-таки имели место. Вот такой удивительный зигзаг претерпели наши представления о пермских флорах России.
Немного тафономии
Рассмотренные выше проблемы органично связаны с вопросом формирования местонахождений, в которых были найдены листья пурсонгий. Все-таки не будем забывать, что внимание палеонтологической общественности к этим растениям было привлечено прежде всего раскопками Амалицкого и открытием знаменитой северодвинской фауны тетрапод с иностранцевиями, скутозаврами, двиниями, котлассиями, карпинскиозаврами и другими палеонтологическими чудесами. Казалось бы, нет такого палеонтолога, изучающего пермскую систему, который так или иначе не соприкоснулся бы с изучением отложений этого возраста, обнажающихся в бассейнах Сухоны и Северной Двины. Однако загадок и противоречий в существующих представлениях об образовании местонахождений наземных растений и тетрапод, расположенных в этом регионе, еще очень много.
Сложность ситуации с родами Glossopteris и Pursongia подчеркивается непростыми, а подчас и драматичными событиями в жизни исследователей, которые соприкоснулись с изучением этих вымерших растений. Выяснилось, что даже ошибочная гипотеза может быть полезна, если она основана на тщательно проработанных фактах. Как тут не вспомнить утверждение И. Канта, что и в самом деле, явления при самом схватывании их суть не что иное, как эмпирический синтез (в пространстве и времени), и, следовательно, даны только в этом синтезе4.
Как этот абзац связан с предыдущим обсуждением и с тафономией местонахождений пурсонгий? Как помнит читатель, Амалицкий именно пурсонгий (которых он считал глоссоптерисами) использовал в качестве одного из доказательств континентальных связей европейской части России (т.е. западных пределов Ангариды) с Гондваной в пермском периоде. Но стало ясно, что пурсонгии и глоссоптерисы — существенно разные растения. Однако дальнейшее изучение пермских флор Ангариды показало, что глоссоптериды в них все-таки есть! Подробнее об этом поговорим чуть позже. Таким образом, в пермском периоде связи между Гондваной и Ангаридой имели место. Вот он, эмпирический синтез Канта, но на палеонтологическом материале.

Общие флористические элементы во флорах Западной Ангариды — Acanthopteridium spinimarginale (а): (1, 2 — прорисовка жилкования; 3 — фотография листа, противоотпечаток которого прорисован на 2) и Гондваны — Ilexoidephyllum permicum (б): 4 — прорисовка жилкования, 5, 6 — фотографии листьев (фиг. 4–6 по [15])
Продолжая эту мысль, неизбежно нужно будет разобраться с тем, как именно, в ходе каких процессов в пространстве и времени, листья пурсонгий сохранились в пермских песчаниках Сухоны и Северной Двины и почему они оказались захороненными вместе с остатками иностранцевий и скутозавров.
Мои представления о тафономии и условиях захоронения листьев пурсонгий в местонахождениях Соколки и Завражье (тех самых, которые изучал Амалицкий на рубеже XIX и XX вв.) сложились благодаря материалам, собранным московскими любителями палеонтологии в начале 1990-х годов и переданным мне для научной обработки. Коллекция была вполне представительной. Начав ее изучение, я обратил внимание на следующее.
Во-первых, и в Соколках, и в Завражье листья пурсонгий часто располагались не строго параллельно плоскости напластования, а были подвернуты и смяты в процессе перемещения вместе с песчано-глинистым материалом, взвешенным в мощном водном потоке. При этом иногда встречались листья, прикреплявшиеся к общему укороченному побегу, т.е. к брахибласту. Во-вторых, в местонахождении Завражье вместе с листьями были найдены и репродуктивные органы пурсонгий, причем представлены они были не отдельными изолированными семеносными дисками, как это обычно бывает, а целыми собраниями — агрегатами из нескольких пельтоидов, прикреплявшихся к общей фертильной оси и формировавших некое подобие шишки — стробила.

Пурсонгия из местонахождения Завражье: а — Pursongia sp., лист, ассоциативно связанный с семеносными органами Peltaspermum polyspermis; б — эпидермально-кутикулярное строение семеносного диска P. polyspermis
Эти наблюдения определенно указывают на то, что листья вместе с репродуктивными органами попали в поток довольно быстро, были сорваны ветром, ливнями или даже самим потоком непосредственно с живого растения и затем, после некоторого переноса, были захоронены вместе с песчанистым материалом. Поэтому важно вспомнить указание Амалицкого на то, что в Соколковской линзе листья пурсонгий встречались преимущественно в верхней части линзы, а скелеты тетрапод (в особенности крупных форм — хищных иностранцевий и растительноядных парейазавров) — в нижней. Это свидетельствует о градационной сегрегации органических остатков в потоке в процессе их перемещения. Тяжелые остатки погружались в нижнюю часть потока, а более легкие оставались в верхней части потока, на плаву.
Твердо убежден в том, что весь поток был одномоментным, и, соответственно, вся толща песков линзы и в Соколках, и в Завражье образовалась за очень короткое время, за считанные дни, а может быть, даже и часы. Скелеты погружались на дно вместе с мягкими тканями, и разложение этих тканей совместно с деятельностью организмов-деструкторов (прежде всего бактерий) привело к образованию мощных конкреций вокруг скелетов и отдельных костей. Именно поэтому мне представляется более правильным сравнивать отложения линз Соколков и Завражья не с русловым или старичным аллювием, а с пролювиальными образованиями, т.е. с отложениями временных, но катастрофических по масштабам водотоков.

Палеофитогеография пермского периода. Растения: 1 — собрание семеносных дисков Peltaspermum; 2 — лист (вайя) пельтаспермового птеридосперма Permocallipteris; 3 — фертильная зона хвощевидного Tchernovia; 4 — латеральный стробил войновскиевого Vojnovskya; 5 — побег войновскиевого с листьями Rufloria; 6 — лист (вайя) птеридосперма из группы гигантоптерид Gigantopteris; 7 — побег хвощевидного Raniganjia; 8 — женский репродуктивный орган глоссоптерид Scutum; 9 — мужской репродуктивный орган глоссоптерид Eretmonia; 10 — лист Belemnopteris. Морфология растений дана по разным авторам, с изменениями и дополнениями. Условные обозначения: а — зона развития бореальной растительности ангарского типа; б — зона развития влаголюбивой тропической растительности катазиатского типа; в — зона развития сухолюбивой тропической растительности еврамерийского типа; г — зона развития нотальной растительности гондванского типа. Красными стрелками показано возможное направление миграции гондванских элементов в западные пределы Ангариды. СТЗ — северная тропическая зона; ЮТЗ — южная тропическая зона
***
Казалось бы, выявленные различия в анатомическом строении классических пурсонгий и глоссоптерид свидетельствуют о том, что гипотеза о связях флор Гондваны и Ангариды себя исчерпала. Но это не так.
Удивительным и красноречивым доказательством связей флор древних континентов Ангариды и Гондваны служит находка в отложениях полдарской свиты (северодвинский ярус верхней перми, согласно современной стратиграфической номенклатуре), которые обнажаются в разрезах Опоки и Никулино (Вологодская обл., Великоустюжский р-н, бассейн р. Сухоны), очень необычного растения, отнесенного к новому роду и виду Acanthopteridium spinimarginale [14]. Это растение очень похоже на вид Ilexoidephyllum permicum, описанный ранее из пермских отложений бассейна р. Параны (Южная Америка) [15]. Единственное и не очень существенное различие — форма верхушки, которая у акантоптеридиума обычно приостренная, а не лопатчатая (или закругленная), как у илексойдефиллума. И кстати, у обоих растений между жилками присутствуют отчетливые и несомненные анастомозы.
Настоящие глоссоптериды были обнаружены в пермских отложениях юго-восточных пределов Ангариды: в Монголии (о новых находках и обзор проблемы см. [16]), Северном Китае [17] и на Дальнем Востоке [18–20]. Это позволяет считать доказанным существование миграционных связей между Гондваной и Ангаридой.
Гондванские элементы могли перемещаться в северном направлении по периферии древнего океана Тетис. Возможность таких миграций была обусловлена флуктуациями климата, при которых растения, приспособленные к относительно суровым сезонно холодным условиям, могли проникать в регионы, занятые низкоширотной растительностью.
О широких миграционных обменах между соседними палеофлористическими областями писали многие палеоботаники. В пермских отложениях Омана (Аравийский п-ов) было обнаружено удивительное смешение гондванских, катазиатских и еврамерийских элементов, которое можно было объяснить, только допустив миграцию исходных видов по периферии Тетиса [21]. О таких же масштабных миграционных обменах между Анаридой и Катазией писали китайские палеоботаники [22]. Остались ли еще какие-то нерешенные вопросы в более чем вековой истории с родами Glossopteris и Pursongia? Да, остались.
Вот самый главный: какие же все-таки репродуктивные органы были связаны с ангарскими (юго-восточно-ангарскими) глоссоптеридами? Такие органы пока неизвестны. Возможно, они уже лежат где-то в лотках с образцами, собранными геологами-съемщиками, но профессиональные палеоботаники до них пока еще просто не добрались. Но возможно эти находки, погребенные в темноте углистых сланцев и песчаников в глубинах Земли, еще ждут своих первооткрывателей. И, кто знает, может быть, кому-то из читателей этой статьи удастся их найти.
Литература
1. Fischer von Waldheim G. N. Nachtrag zu Hrn. Major von Qualens geognostischen Beyträgen zur Kenntniss des westlichen Urals // Bulletin de la Société impériale des naturalistes de Moscou. 1840; (IV): 488–494.
2. Mercklin C. Verzeichnis aller in Russland bis jetzt (November 1852) aufgefundener beschriebener, unbeschriebener oder zweifelhafter fossiler Pflanzen // Mémoires physique et chimie de l’Académie de Sciences. 1852. 1: 497–504.
3. Suess E. The face of the Earth. Oxford, 1904.
4. Нейбург М. Ф. Опыт фитостратиграфического сопоставления верхнепалеозойских отложений Ангариды и Гондваны (Индия) // Вопросы геологии Азии. 1954; 1: 767–797.
5. Залесский М. Д. Палеозойская флора Ангарской серии: Атлас. Пг., 1918 (Труды Геологического комитета. Новая серия; Вып. 174).
6. Залесский М. Д. О Cordaites aequalis Göppert из Сибири и о тождестве его с Neggerathiopsis hislopii Bunbury sp. флоры Гондваны. СПб., 1912. (Труды Геологического комитета. Новая серия; Вып. 86).
7. Залесский М. Д. Гондванская флора бассейна реки Печоры. 1: Река Адзьва. Екатеринбург, 1914; 55–85 (Записки Уральского общества любителей естествознания. Т. 33).
8. Залесский М. Д. Пермская флора Уральских пределов Ангариды: Атлас. Л., 1927 (Труды Геологического комитета. Новая серия; Вып. 176).
9. Залесский М. Д. Распространение ископаемой флоры, родственной гондванской, в пределах северной части Евразии // Известия Академии наук. VII серия. Отделение математических и естественных наук. 1930; (9): 913–930.
10. Мейен С. В. О роде Zamiopteris Schmalhausen и его соотношении с некоторыми смежными родами. Птеридоспермы позднего палеозоя и мезозоя. М., 1969; 85–104. (Труды Геологического института РАН, Вып. 190).
11. Наугольных С. В. Вязниковская флора и природа пермо-триасового вымирания. Причинно-следственные связи и факторы глобальных биосферных перестроек в фанерозое. М., 2006; 42–71. (Труды Геологического института РАН, Вып. 580).
12. Naugolnykh S. V. A new peltaspermaceous pteridosperm from the Upper Permian of the Russian platform // The Palaeobotanist. 2001; 50: 189–205.
13. Naugolnykh S. V. Basic reproductive strategies of glossopterids: supraidioadaptive divergence from the viewpoint of a non-Gondwana palaeobotanist // The Palaeobotanist. 2016; 65: 1–17.
14. Наугольных С. В., Арефьев М. П. Новое загадочное растение из верхней перми Субангариды // Доклады международного симпозиума «Верхнепермские стратотипы Поволжья». М., 1999; 283–295.
15. Rohn R., Rösler O. Folhas denteadas da Farmacao Rio do Rasto (Bacia do Parana, Permiana Superior) e seu possivel Significade paleoclimatico // Boletin de Instituto Geologico. 1989; (7): 127–137.
16. Naugolnykh S. V., Uranbileg L. A new discovery of Glossopteris in southeastern Mongolia as an argument for distant migration of Gondwanan plants // Journal of Asian Earth Sciences. 2018; 154: 142–148.
17. Yang T., Naugolnykh S. V., Sun G. A new representative of Neocalamites Halle from the Upper Permian of Northeastern China (Jiefangcun Formation) // Paleontological Journal. 2011; 45(3): 335–346.
18. Зимина В. Г. Первая находка Glossopteris и Gangamopteris в пермских отложениях Южного Приморья // Вопросы геологии и рудоносности Дальнего Востока. Тезисы докладов на юбилейной сессии Дальневосточного геологического института ДВ филиала СО АН СССР, посвященной пятилетию института. Владивосток, 1965; 49–50.
19. Зимина В. Г. О Glossopteris и Gangamopteris из пермских отложений Южного Приморья // Палеонтологический журнал. 1967; (2): 113–121.
20. Зимина В. Г. Флора ранней и начала поздней перми Южного Приморья. М., 1977.
21. Broutin J., Roger J., Platel J. P. et al. The Permian Pangea. Phytogeographic implications of new paleontological discoveries in Oman (Arabian Peninsula) // Comptes rendus de l’Académie des Sciences, Paris. Ser. IIa. 1995; 321: 1069–1086.
22. Wang Z.-Q. Past global floristic changes: the Permian great Eurasian floral interchange // Palaeontology. 1996; 39(1): 189–217.
23. Schmalhausen J. Beitrage zur Jura-Flora Russlands. St.-Peterbourg, 1879; (Mémoires de l’Académie impériale des sciences de St.-Peterbourg. VII Serie. T. XXVII, №4).
24. Zalessky M. D. Flores permiennes du la plaine Russe, de l’Oural et du bassin de Kousnetzk et les correlations des dépôts qui les contiennent // Problems of Paleontology. V. II–III. Moscow, 1937; 9–35.
25. Залесский М. Д. О новых ископаемых растениях антраколитовой системы Кузнецкого бассейна. I // Известия Академии наук. VII серия. Отделение математических и естественных наук. 1933; (8): 1213–1258.
26. Нейбург М. Ф. Верхнепалеозойская флора Кузнецкого бассейна. М; Л., 1948.
1 Голосницкий Л. П. Путешествие в прошлое. М., 1957. С. 60.
2 Сорокина М. Ю. Наука и Третий рейх: борьба за ресурсы // Природа. 2003. №8. С. 73–80.
3 Наугольных С. В. Символ Гондваны // Природа. 2012. №12. С. 49–59.
4 Кант И. Критика чистого разума. Минск, 1998. С. 534.


















Листья пурсонгий из работ М. Д. Залесского: а — Pursongia amalitzkii, местонахождение Соколки, Малая Северная Двина [24, fig. 2]; б — Pursongia amalitzkii, левый берег р. Сакмары у д. Кульчумкино, ст. Маячная [24, fig. 1]; в — Pursongia asiatica, Кузбасс, д. Ишановка, кровля пласта Нового [25, фиг. 23; 26, рис. 24]