Живые индикаторы метановых выходов и гидротерм
Елена Крылова, Евгений Колпаков
«Природа» №6, 2019
 Елена Михайловна Крылова — кандидат биологических наук, старший научный сотрудник лаборатории донной фауны океана Института океанологии имени П. П. Ширшова РАН. Принимала участие в российских и международных океанических научных рейсах. Область научных интересов — фауна восстановительных биотопов, систематика, морфология, распространение и эволюция глубоководных двустворчатых моллюсков. |
 Евгений Викторович Колпаков — научный сотрудник лаборатории бентоса Тихоокеанского филиала Всероссийского научно-исследовательского института рыбного хозяйства и океанографии. Область научных интересов — фауна, распространение и экология двустворчатых моллюсков дальневосточных морей. |
В число задач 82-го рейса научно-исследовательского судна «Академик М. А. Лаврентьев» были включены поиск и описание восстановительных биотопов на Корякском склоне Берингова моря. О существовании в этом районе восстановительных условий свидетельствовало присутствие в траловых уловах научно-исследовательских судов живых двустворчатых моллюсков плиокардиин и створок их раковин [1, 2]. В июле 2018 г. с помощью телеуправляемого необитаемого подводного аппарата Comanche 18 («Команч») было найдено самое северное местообитание плиокардиин в Мировом океане, отобраны биологические пробы, получены редкие фото- и видеоматериалы [3].
Для того чтобы объяснить читателю, почему летом 2018 г. места находок плиокардиин стали «путеводной нитью» наших изысканий и какими особенностями этих двустворок обусловлена их приуроченность к районам гидротерм и высачиваний углеводородов, перевернем назад несколько страниц истории изучения этих глубоководных моллюсков и вернемся в последнюю треть XX в.
Загадочные гигантские двустворки
Двустворки подсемейства плиокардиин (Bivalvia: Vesicomyidae) — одни из самых ярких представителей глубоководных сообществ, развивающихся на основе хемосинтеза в так называемых восстановительных условиях. В течение почти 100 лет, прошедших со времени описания первых видов этого подсемейства до открытия глубоководных восстановительных биотопов на дне океана в 1977 г., эти моллюски оставались самой таинственной и малоизученной группой глубоководных двустворок. В сборах экспедиций они встречались крайне редко и обычно на глубинах свыше 1000 м. Как правило, размеры глубоководных двустворок невелики: моллюски длиной 3 см уже считаются гигантами. Однако у подавляющего большинства плиокардиин длина тела превышает 3,5 см и в среднем составляет 8–10 см. Им же принадлежит рекорд среди глубоководных двустворок: длина отдельных экземпляров достигает 30 см (!). Столь значительные размеры тела при довольно неблагоприятных трофических условиях нижней батиали и абиссали — поистине парадокс! И он требовал объяснения.
Поскольку при отборе проб попадались, в основном, пустые створки моллюсков, ученые предполагали, что эти фрагменты могли быть снесены течениями с шельфа. Однако в мелководных местообитаниях загадочных двустворок тоже не находили...
В рамках существовавшей до 1977 г. парадигмы, согласно которой все органическое вещество Мирового океана образуется в его поверхностных горизонтах или поступает с суши, было невозможно разгадать природу существования в глубоководье (т.е. в условиях хронического недостатка питательных веществ) крупноразмерных животных, формирующих популяции высокой плотности. Попытка объяснить процветание гигантов на глубинах наличием альтернативного источника органики — хемосимбиотрофных бактерий, которые живут в клетках их тела — в то время граничила бы с фантастикой.

В 1977 г., когда на глубинах около 3000 м в районе Галапагосского рифта с помощью американского подводного обитаемого аппарата «Алвин» были обнаружены выходы горячих растворов, обогащенных сероводородом, солями металлов и водородом, начался новый этап в истории изучения плиокардиин. В непосредственной близости от выходов гидротермальных флюидов процветали богатые сообщества необычных животных, питающихся, как было показано позднее, органическим веществом хемосинтетического происхождения. Одним из доминирующих видов сообществ была гигантская двустворка неизвестного науке вида (рис. 1), который получил говорящее название magnifica [4]. Этот вид плиокардиин стал символом эпохального открытия гидротермальных систем.
Вслед за гидротермами были открыты холодные углеводородные высачивания и связанная с ними фауна на дне Мексиканского залива и у берегов штата Орегон в северо-восточной части Тихого океана. Затем последовали многочисленные сообщения о новых находках «оазисов» на дне океана. В отечественных исследованиях восстановительных биотопов ключевую роль сыграли подводные обитаемые аппараты «Пайсис», «Мир-1» и «Мир-2» [5]. Повышенные концентрации метана и сульфидов были отмечены не только в районах гидротерм и холодных высачиваний, но также и в местах скопления разлагающейся органики, например, около затонувших скелетов китов. Практически в каждом из этих биотопов встречались, а часто и доминировали двустворки плиокардиины.
В настоящее время в подсемействе Pliocardiinae описано более 110 ныне живущих видов, распространенных в восстановительных биотопах по всему Мировому океану на глубинах от 100 до 6809 м [6]. Плиокардиины вошли в число самых изученных глубоководных групп двустворчатых моллюсков и стали модельным объектом для изучения физиологии, биохимии, популяционной генетики и эволюции хемосимбиотрофных организмов. Практически весь материал для этих исследований был получен с помощью подводных аппаратов, без использования которых изучение восстановительных сообществ Мирового океана было бы невозможно [5].
Бактериальная симбиотрофия
Плиокардиины — наиболее успешная группа двустворок восстановительных биотопов, по числу видов они лидируют среди двустворчатых моллюсков районов углеводородных высачиваний. Биомасса плиокардиин может превышать 30 кг/м2, что сопоставимо с биомассой мидиевых банок на мелководье. Успех группы обеспечил целый комплекс морфологических, физиологических и биохимических адаптаций, главной из которых стало развитие внутриклеточного симбиоза с хемолитоавтотрофными бактериями.
Все до сих пор изученные виды плиокардиин живут в обязательном симбиозе с сульфид-окисляющими (тиотрофными) гамма-протеобактериями [7]. Эти бактерии содержатся в специализированных клетках эпителия жабр, называемых бактериоцитами. Моллюск снабжает симбионтов необходимыми для их функционирования сульфидами, кислородом и углекислым газом. А бактерии, используя энергию окисления сульфидсодержащих восстановленных соединений, синтезируют органическое вещество из неорганического углерода, за счет которого и живет моллюск [8]. Предполагают, что моллюск получает питательные вещества посредством лизиса бактериальных клеток, в результате чего органические молекулы попадают в цитоплазму бактериоцитов, а далее в гемолимфу. Молекулярный анализ показал, что по сравнению со свободноживущими бактериями геномы симбиотических бактерий имеют значительно меньшие размеры. Так, геном симбионта Calyptogena magnifica в два раза меньше геномов их ближайших свободноживущих родственников [9]. Это свидетельствует о значительных перестройках физиологии участников симбиотического взаимодействия. Основной путь «заражения» плиокардиин бактериальными симбионтами — так называемый вертикальный: бактерии передаются от материнских особей к следующим поколениям моллюсков через яйцеклетки.
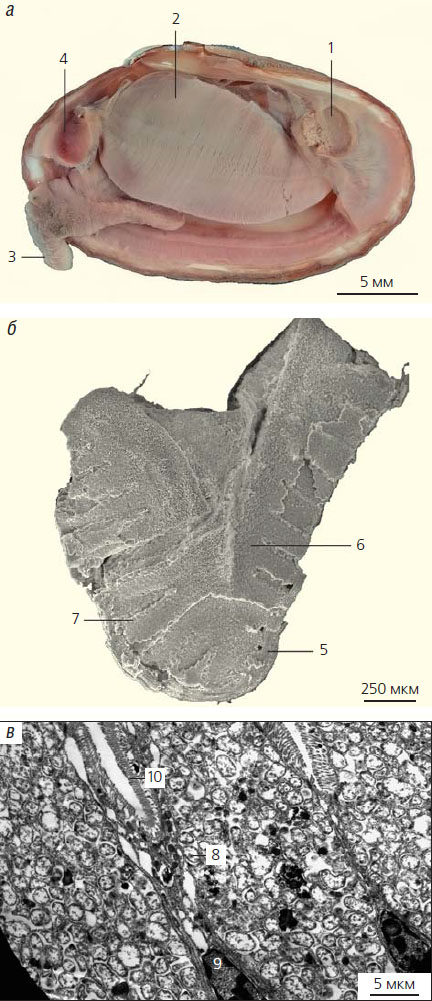
Рис. 2. Строение плиокардиины: а — тело Calyptogena pacifica (левая створка удалена), 1 — задний мускул-замыкатель, 2 — левая жабра, 3 — нога, 4 — передний мускул-замыкатель; б — внутреннее строение жабры (сканирующий электронный микроскоп), 5 — жаберный филамент; 6 — септа, состоящая из бактериоцитов и заполняющая пространство между филаментами, 7 — отдельные бактериоциты, различаемые в септе; в — жаберный эпителий плиокардиины, в котором видны специализированные клетки бактериоциты, содержащие многочисленные бактерии, и узкие промежуточные клетки с ядрами у основания бактериоцитов (просвечивающий электронный микроскоп), 8 — бактерии, 9 — ядро, 10 — микровилли
Изначально на поверхности каждой яйцеклетки имеется около 400 бактерий, которые в процессе развития и дифференциации тканей и органов моллюска размножаются и занимают внутриклеточное положение в тканях жабр [10]. В целом, бактерии видоспецифичны для плиокардиин (каждый вид моллюсков содержит свой вид бактерий). Однако описано несколько случаев, когда в тканях моллюска помимо обычных его симбионтов присутствовало небольшое число бактерий, характерных для живущего поблизости вида плиокардиин [11]. Механизм такой передачи симбионтов еще не известен. Установление симбиотических отношений с тиотрофными бактериями дало возможность плиокардиинам широко освоить глубоководные восстановительные биотопы.
Вовлечение моллюсков в симбиотические отношения с бактериями вызвало значительные анатомические, биохимические и физиологические перестройки их организма. Поскольку жабры превратились в орган-инкубатор для бактерий, у каждого жаберного филамента появилась дополнительная часть — так называемая септа, состоящая из клеток-бактериоцитов, заполненных бактериями (рис. 2). Пищеварительная система, напротив, уменьшилась. Кишка самых приспособленных к симбиотрофии моллюсков превратилась в тонкую, лишенную петлей трубку. Пищеварительная камера («желудок») также мала и отличается упрощенной внутренней структурой.
В отличие от подавляющего большинства двустворчатых моллюсков, у плиокардиин в гемолимфе присутствует внутриклеточный гемоглобин, отчего она окрашена в красный цвет. Более того, виды плиокардиин могут различаться по структуре и числу видов гемоглобина, имеющего у этих животных в среднем очень высокое сродство к кислороду (хотя оно варьирует у разных видов) [12]. Способность гемоглобина эффективно связывать кислород имеет особое значение в восстановительных местообитаниях.
Живая летопись тектонической активности на дне океана
Поскольку плиокардиины живут в обязательном симбиозе с тиотрофными бактериями, повышенное содержание сульфидов в осадке — непременное условие их существования. Источник повышенной концентрации сульфидов может быть любым. В случае гидротерм, восстановленные соединения, включая H2S и сульфиды металлов, присутствуют в горячих растворах, выделяющихся из трещин океанической коры. В районах холодных углеводородных выходов присутствие в осадках сульфидов — результат анаэробного окисления метана. В целом для подсемейства нижняя граница необходимой концентрации сульфидов в осадках составляет примерно 100 мкмоль.
Мы составили карту всех местообитаний плиокардиин, включая литературные и неопубликованные оригинальные данные (рис. 3). 33 вида плиокардиин были описаны до открытия феномена восстановительных сообществ и собраны с помощью тралов «вслепую». Оказалось, что все старые находки расположены в районах, где впоследствии обнаружены углеводородные или гидротермальные выходы.

Рис. 3. Карта распространения двустворок плиокардиин (Bivalvia: Vesicomyidae)
Открытие донных восстановительных биотопов и связанных с ними специализированных сообществ на Корякском склоне стало одной из наиболее ярких научных удач экспедиции. Было собрано много материала, требующего длительной обработки и анализа, так что детальное обсуждение результатов и основные выводы еще впереди. Но уже сейчас можно с уверенностью сказать: открыты самые северные в бассейне Тихого океана восстановительные биотопы. Кроме того, это самая северная в мире область, где обнаружены современные плиокардиины. Ранее в Беринговом море восстановительные условия и связанные с ними сообщества были отмечены только на вулкане Пийпа, расположенном в юго-западной части Берингова моря на склоне Командорских островов*. Тогда исследования проводились с применением обитаемых глубоководных аппаратов Института океанологии имени П. П. Ширшова РАН [13].
Пока на Корякском склоне встречены два вида плиокардиин (Calyptogena pacifica и Phreagena soyoae). C. pacifica обнаружена на глубинах от 400 до 695 м, причем на 650 м вид образовывал очень плотные поселения (рис. 4). В тех же поселениях были встречены единичные экземпляры P. soyoae.

Рис. 4. Популяция Calyptogena pacifica в природе. Корякский склон, 660 м, биотоп метановых выходов. Расстояние между лазерными точками — 5 см. Фото предоставлено Национальным научным центром морской биологии ДВО РАН
Еще до того как состоялись наши находки, было известно, что оба вида, несмотря на узкоспецифические требования к среде, имеют протяженные амфипацифические ареалы — т.е. обитают и в Западной, и в Восточной Пацифике. Молекулярные данные указывали на высокий уровень генетического сходства популяций из удаленных частей ареалов [14]. Однако механизм поддержания генетического сходства на всем протяжении области распространения был непонятен. О популяциях, которые могли бы обеспечивать связь между различными частями ареала, известно не было.
Новые находки плиокардиин в Беринговом море значительно уменьшают расстояние между западными и восточными частями ареалов видов. Кроме того, полученные в ходе нашей экспедиции данные о количественном распространении метана в толще воды на Корякском склоне** указывают на более широкое, чем известно в настоящее время, распространение восстановительных условий, которые могут поддерживать еще не обнаруженные специализированные сообщества. Получается, что западная и восточная части ареалов связываются через северный «мост» из уже открытых и предполагаемых восстановительных биотопов. Заметим: в северной части Тихого океана более трети видов плиокардиин имеют амфиокеанический ареал, что значительно больше, чем в Атлантике, где западный и восточный континентальные склоны расположены ближе друг к другу, но не связаны цепочкой подходящих для плиокардиин биотопов [2].
Околоконтинентальные восстановительные биотопы в северной Пацифике располагаются в батиали (от 300 до 3000 м), обеспечивая пути распространения видам, живущим именно на этих глубинах.
Интересно, что сходство фаун плиокардиин западной и восточной частей Северной Пацифики проявляется не только в большой доле общих видов, но и в наличии близкородственных сестринских — так называемых видов-двойников. В отличие от амфипацифических видов, «двойники» обитают преимущественно в абиссали, т.е. глубже 3000 м. Возможно, виды-двойники появились у западного и восточного континентальных склонов Пацифики в результате более ранних миграций, которые невозможны в современных условиях. Время дивергенции видов двойников, имеющих общего предка, определено на основе молекулярных данных [15] и приходится на конец миоцена — начало плиоцена. Известно, что именно в этот период в Северной Пацифике возросла тектоническая активность, что, в частности, привело к открытию Берингова пролива около 5,4 млн лет назад [16]. С тектонической активностью на дне океана связано появление восстановительных биотопов, что могло способствовать усилению транспацифических миграций абиссальных плиокардиин.
Прочная связь распространения плиокардиин с районами повышенной концентрации сульфидов делает их удобными и надежными индикаторами восстановительных условий на дне океана. Данные о нахождениях плиокардиин дают возможность выявлять области углеводородных высачиваний и гидротермальных выходов, а также реконструировать восстановительные условия, существовавшие в прошлом.
На карте распространения плиокардиин часть современных находок совпадает с областями, в которых восстановительные биотопы пока не известны, однако их присутствие можно прогнозировать с очень высокой степенью вероятности (см. рис. 3). Так, в частности, мы можем ожидать наличие восстановительных условий в абиссали Бискайского залива (восточная часть Атлантического океана) и на склоне о. Мадагаскар (западная часть Индийского океана). Анализ ареалов моллюсков однозначно свидетельствует о том, что восстановительные условия в Мировом океане распространены шире, чем это предполагается сейчас.
Мы благодарим В. А. Надточего (Тихоокеанский филиал Всероссийского научно-исследовательского института рыбного хозяйства и океанографии (ТИНРО)) за материалы плиокардиин с Корякского склона Берингова моря, собранные в экспедициях ТИНРО. Е. М. Крылова глубоко благодарна сотрудникам и руководству Национального научного центра морской биологии имени А. В. Жирмунского ДВО РАН за приглашение принять участие в экспедиции, всему научному составу экспедиции, капитану В. Б. Птушкину и команде судна «Академик М. А. Лаврентьев», пилотам и техникам телеуправляемого необитаемого подводного аппарата «Команч» за высокопрофессиональную работу.
Работа Е. М. Крыловой частично поддержана грантом Российского фонда фундаментальных исследований (проект 18-05-60228).
Литература
1. Данилин Д. Д. Двустворчатые моллюски как потенциальные индикаторы районов гидротермальной активности // Материалы конференции, посвященной Дню вулканолога «Вулканизм и связанные с ним процессы». Петропавловск-Камчатский, 2013; 291–294.
2. Krylova E. M., Kolpakov E. V., Sharina S. N. et al. Distribution patterns of chemosymbiotic bivalves of the subfamily Pliocardiinae (Bivalvia: Vesicomyidae) from the North-West Pacific // 15-th International deep-sea biological symposium, Monterey (USA). 2018; 32–33.
3. Галкин С. В., Мордухович В. В., Крылова Е. М. и др. Комплексные исследования экосистем гидротермальных выходов и холодных высачиваний в Беринговом море (82-й рейс НИС «Академик М. А. Лаврентьев») // Океанология. 2019; 4 (в печати).
4. Boss K. J., Turner R. D. The giant white clam from the Galapagos rift, Calyptogena magnifica species novum // Malacologia. 1980; 20: 161–194.
5. Сагалевич А. М. Роль визуальных наблюдений в глубоководных исследованиях океана // Природа. 2018; 8: 58–66. DOI: 10.31857/S0032874X0000489-6.
6. Krylova E. M., Sahling H. Vesicomyidae (Bivalvia): current taxonomy and distribution // PLoS One. 2010; 5(4): e9957. DOI: 10.1371/journal.pone.0009957.
7. Dubilier N., Bergin C., Lot C. Symbiotic diversity in marine animals: The art of harnessing chemosynthesis // Nature Reviews Microbiology. 2008; 6: 725–740. DOI: 10.1038/nrmicro1992.
8. Cavanaugh C. M. Symbiotic chemoautotrophic bacteria in marine invertebrates from sulphide-rich habitats // Nature. 1983; 302: 58–61. DOI: 10.1038/302058a0.
9. Newton I. L. G., Woyke T., Auchtung T. A. et al. The Calyptogena magnifica chemoautotrophic symbiont genome // Science. 2007; 315: 998–1000. DOI: 10.1126/science.1138438.
10. Ikuta T., Igawa K., Tame A. et al. Surfing the vegetal pole in a small population: extracellular vertical transmission of an «intracellular» deep-sea clam symbiont // Royal Society Open Science. 2016; 3: 160130. DOI: 10.1098/rsos.160130.
11. Decker C., Olu K., Arnaud-Haond S. et al. Physical proximity may promot lateral acquisition of bacterial symbionts in vesicomyid clams // Public Library of Science One. 2013; 8: e64830—64812. DOI: 10.1371/journal.pone.0064830.
12. Decker C., Zorn N., Potier N.,et al. Globin’s structure and function in vesicomyid bivalves from the Gulf of Guinea cold seeps as an adaptation to life in reduced sediments // Physiological and Biochemical Zoology. 2014; 87: 855–869. DOI: 10.1086/678131.
13. Сагалевич А. М., Торохов П. В., Галкин С. В. и др. Гидротермальные проявления подводного вулкана Пийпа (Берингово море) // Изв. РАН. Сер. геолог. 1992; 9: 104–114.
14. Audzijonyte A., Krylova E. M., Sahling H., Vrijenhoek R. C. Molecular taxonomy reveals broad trans-oceanic distributions and high species diversity of deep-sea clams (Bivalvia: Vesicomyidae: Pliocardiinae) in chemosynthetic environments // Systematics and Biodiversity. 2012; 10: 403–415. DOI: 10.1080/14772000.2012.744112.
15. Johnson S. B., Krylova E. M., Audzijonyte A. et al. Phylogeny and origins of chemosynthetic vesicomyid clams // Systematics and Biodiversity. 2017; 1–15. DOI: 10.1080/14772000.2016.1252438.
16. Gladenkov A. Y., Gladenkov Y. B. 2004. Onset of connections between the Pacific and Arctic Oceans through the Bering Strait in the Neogen // Stratigr. Geol. Correl. 2004; 12: 175–187.
* Островной склон — подводный склон острова, лежащего вне шельфовой зоны.
** См. статью Н. С. Полоника (с. 36).













Рис. 1. Популяция Calyptogena magnifica в природе. Калифорнийский залив, 2290 м, гидротермальный биотоп. Длина раковин на снимке около 20 см. Фото любезно предоставлено MBARI (Исследовательский институт аквариума Монтерея) с разрешения Р. Врайенхука