Новости о древних людях
Михаил Гельфанд
«Природа» №11, 2016
 Об автореМихаил Сергеевич Гельфанд — доктор биологических наук, профессор, член Европейской академии, заместитель директора Института проблем передачи информации им. А. А. Харкевича РАН, профессор факультета биоинженерии и биоинформатики Московского государственного университета им. М. В. Ломоносова. Область научных интересов — биоинформатика, молекулярная эволюция, системная биология, сравнительная и функциональная геномика, метагеномика. |
Новые результаты исследований, посвященных ДНК древних людей, на часть вопросов, которые мы обсуждали почти год назад [1], отвечают, какие-то из них оставляют открытыми, а некоторые — ставят заново.
Были проанализированы денисовские фрагменты в геномах австранезийцев и кроманьонские — в геномах неандертальцев. В геномах африканцев были найдены некроманьонские фрагменты, что указывает на гибридизацию с какими-то неизвестными представителями рода Homo. Продолжились исследования отдельных групп генов. Надо сказать, что картина взаимоотношений между ветвями человечества стала более подробной, но и более запутанной. Кроме того, некоторые исследования, убедительные сами по себе, друг другу противоречат.
Древние геномы
Была определена последовательность Y-хромосомы (точнее, ее белок-кодирующих фрагментов) неандертальца из испанской пещеры Эль-Сидрон. Это первая секвенированная древняя Y-хромосома [2]. Ее эволюционная история не имеет неожиданностей — она более или менее совпадает с историей соматических хромосом. Зато удалось выяснить, продукты каких генов могли способствовать репродуктивной изоляции неандертальцев: три белка, в генах которых наблюдаются различия между неандертальцами и кроманьонцами, по-видимому, могли вызывать потенциальный иммунный конфликт между матерью и сыном. Это согласуется с классическим правилом Холдейна: если при скрещивании разных подвидов или рас жизнеспособность потомства зависит от пола, более редким (или вообще отсутствующим) будет гетерогаметный пол, то есть у млекопитающих — самцы [3]. Таким образом, в браках между неандертальцами и кроманьонцами, возможно, не было сыновей и почти наверняка — внуков по мужской линии. Авторы статьи заключают, что у потомков таких скрещиваний, т. е. современных европейцев и азиатов, не была обнаружена неандертальская Y-хромосома, однако это утверждение кажется излишним. Действительно, никого ведь не удивляет, что у современных людей не было обнаружено неандертальских митохондриальных геномов, передающихся, наоборот, строго по материнской линии. В связи с этой работой можно вспомнить и более раннее наблюдение: неандертальские варианты в геномах современных людей относительно редки в Х-хромосомах и полностью отсутствуют в генах, экспрессирующихся в семенниках, что говорит о повышенной мужской стерильности потомков смешанных пар [4].
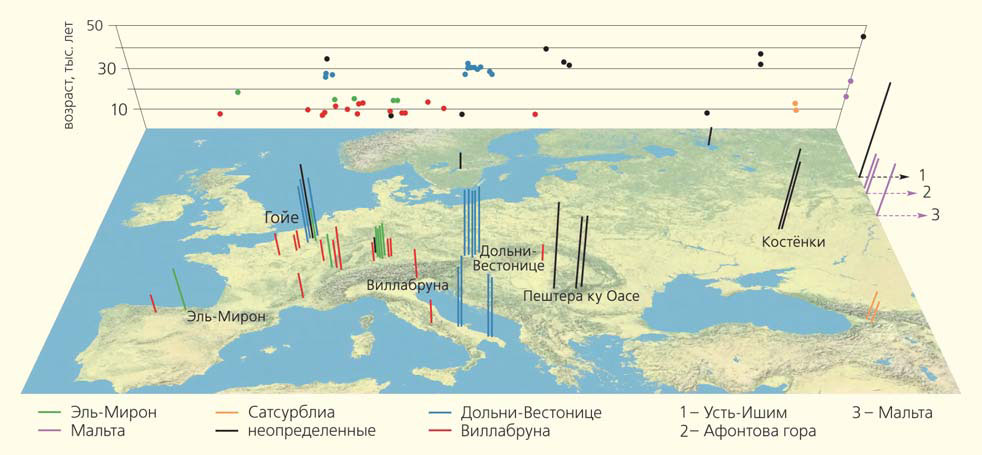
Многие неандертальские аллели, по-видимому, были в той или иной степени вредны, так как в 51 исследованном геноме древних евроазиатов возрастом от 7 до 45 тыс. лет доля неандертальских вариантов постепенно сокращалась [5] (рис. 1, 2). Впрочем, отмечено, что частота неандертальских фрагментов падает вблизи границ областей структурных перестроек генома — видимо, последние препятствуют рекомбинации [6].
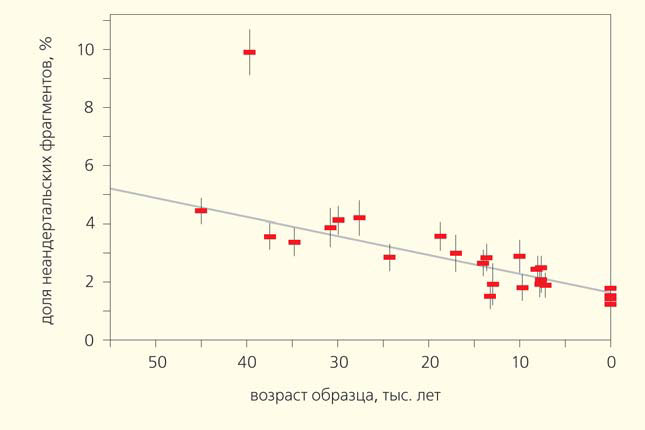
Рис. 2. Зависимость доли неандертальских фрагментов в геномах древних кроманьонцев от возраста образца ДНК. Выделяется геном представителя тупиковой ветви, останки которого были найдены в румынской пещере Пештера-ку-Оасе, — гибридизация произошла за 4–6 поколений до него. В качестве современных взяты геномы азиатов
Секвенирование еще двух денисовских геномов показало, что, судя по числу замен, самый древний геном старше самого молодого на 60 тыс. лет [7]. К сожалению, качество ядерных геномов не позволило провести более детальные исследования, например, чтобы понять, какой из геномов ближе к денисовскому предку австранезийцев. Был также секвенирован ядерный геном человека из испанской пещеры Сима де лос Уэсос возрастом около 430 тыс. лет, относящийся к среднему плейстоцену [8]. Напомним, что этот человек оказался ближайшим родственником денисовцев по материнской линии: именно на его митохондриальный геном больше всего похож митохондриальный геном денисовца. Оба отделились от ветви неандертальцев и кроманьонцев примерно миллион лет назад. Однако два ядерных генома из пещеры Сима де лос Уэсос говорят о другом: они существенно больше похожи на геном алтайского неандертальца, чем на геномы денисовца и кроманьонца. Получается, интрогрессия митохондриальной ДНК могла случиться не от неизвестных представителей рода Homo (обсуждался даже H. erectus) к людям из Сима де лос Уэсос и из Денисовой пещеры (независимо?), а от африканских кроманьонцев к неандертальцам, причем первоначальные неандертальские митохондриальные геномы в позднем плейстоцене уже не сохранились. Расхождение ветвей кроманьонцев с неандертальцами и денисовцами при этом сдвигается в более ранний период, чем полагали до сих пор, — по-видимому, это произошло 765–550 тыс. лет назад [9].
Анализ уже секвенированных последовательностей — полных геномов алтайских неандертальца и денисовца и 21-й хромосомы неандертальцев из испанской пещеры Эль Сидрон и хорватской пещеры Виндия — показал наличие кроманьонских фрагментов в геноме алтайского неандертальца, но не в трех других [10]. Анализ последовательностей и моделирование популяционной истории показывает, что эти фрагменты появились в результате гибридизации, случившейся около 100 тыс. лет назад. Если это верно, то их источник — неизвестная ветвь кроманьонцев, вышедших из Африки задолго до общих предков современных европейцев и азиатов и гибридизовавшихся с одной из групп неандертальцев. Интересно, что один из кроманьонских фрагментов в геноме алтайского неандертальца включает ген FOXP2, отвечающий за развитие речи. В геномах современных людей эта область никогда не содержит неандертальских вариантов, а стало быть, для современного человека неандертальские варианты этой области вредны, в то время как для неандертальца кроманьонский вариант, похоже, оказался полезным. Кроме того, было подтверждено наличие в денисовском геноме фрагментов, происходящих от представителя неизвестной ветви рода Homo (рис. 3, слева).
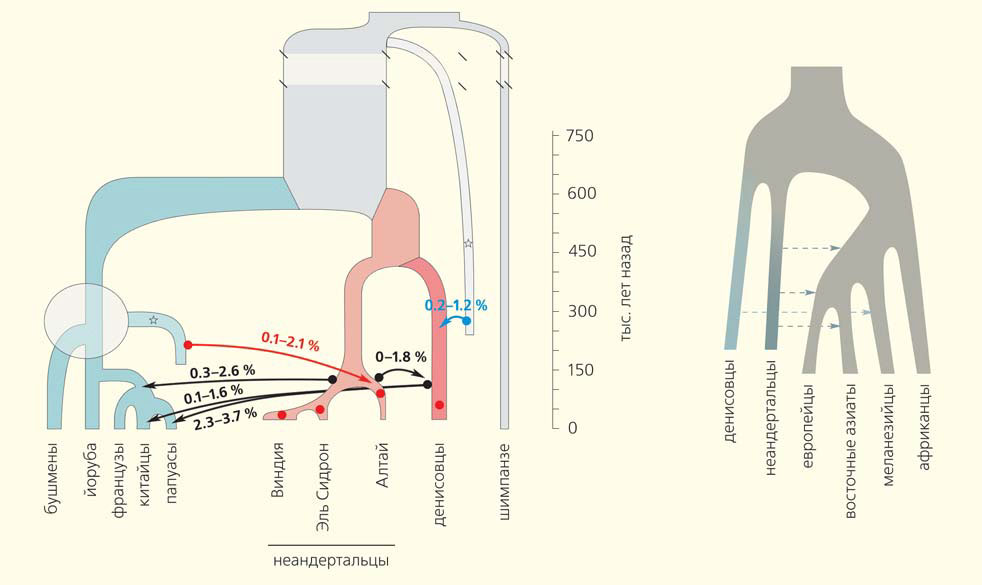
Рис. 3. Реконструкция событий гибридизации [10, 11]. Кружком (слева) обведено место, соответствующее ответвлению людей (ветвь помечена голубой звездочкой), гибридизовавшихся с предками алтайских неандертальцев, — либо вскоре после первого расхождения современных популяций (ответвления койсанских народов), либо еще раньше. Числа над стрелками обозначают среднюю долю фрагментов, появившихся в геномах представителей популяции-реципиента в результате каждого события гибридизации. Серая звездочка — неизвестная древняя ветвь. Различия в порядке ответвления европейцев, китайцев и папуасов на рисунках слева и справа отражают противоречия в цитируемых статьях
Современные геномы
Как и ожидалось, сразу несколькими группами был проведен анализ денисовских и неандертальских фрагментов в геномах сотен современных людей, в том числе десятков меланезийцев [11, 12].
В первой из этих работ было реконструировано в общей сложности 1,34 млрд нуклеотидов (примерно треть) неандертальского генома и 303 млн нуклеотидов (примерно одна десятая часть) денисовского [11]. Авторы постулировали, что имели место как минимум три волны гибридизации предков современных людей с неандертальцами: общего предка европейцев и азиатов (включая меланезийцев), предка европейцев и восточных азиатов (после ответвления меланезийцев) и предка восточных азиатов (рис. 3, справа). Стало уже традиционным определять области с повышенной частотой неандертальских вариантов и те, в которых следов древних геномов нет вовсе. Так вот, последние содержат много старых знакомых: гены, экспрессирующиеся в некоторых областях мозга, гены, связанные с аутизмом, и, конечно, FOXP2, ассоциирующийся с развитием речи. В геномах меланезийцев нашлась 21 область, в которой древние варианты могли быть адаптивными: 14 — неандертальского происхождения, три — денисовского, три — неясного (невозможно различить) и одна область, в которой с повышенной частотой встречаются и неандертальские, и денисовские варианты. И опять эти области содержали гены различных метаболических систем, в том числе метаболизма липидов, и гены, связанные с иммунной системой.
Во второй работе был проведен анализ длин фрагментов денисовского и неандертальского происхождения из 257 современных геномов (представителей 120 популяций) [12]. На его основании было показано, что гибридизация предков современных австранезийцев с денисовцами случилась позднее, чем с неандертальцами, так как денисовские фрагменты оказались длиннее. Это укладывается в общепринятый сценарий, согласно которому гибридизация с неандертальцем произошла у общего предка всех евроазиатов, а с денисовцем — у предка австранезийцев. Как и в ряде других работ, доля денисовских фрагментов в геномах восточных азиатов оказалась выше, чем можно было предположить, исходя из более поздних взаимодействий с жителями Океании, а значит, следует предположить независимую гибридизацию предков восточных азиатов с денисовцами. Анализ областей, в которых никогда не находили древние аллели, показал преобладание областей Х-хромосомных локусов, генов, экспрессирующихся в семенниках, и генов, находящихся под действием положительного отбора в линии современного человека. Все это согласуется с другими наблюдениями и, в частности, указывает на пониженную фертильность мужского потомства смешанных пар.
Были продолжены исследования по поиску следов геномов древних людей в геномах современных африканцев: кочевых западно-африканских пигмеев ака (биака) и бака, живущих за счет охоты и собирательства на границах Центрально-Африканской Республики, Конго, Камеруна и Габона (рис. 4). У них обнаружилось 265 фрагментов общей длиной примерно 20 млн нуклеотидов (две трети процента от размера генома), происходящих от генома, разошедшегося с геномом H. sapiens около миллиона лет назад [13]. Время гибридизации было определено с очень малой точностью: наилучшая оценка составляет 30 тыс. лет назад, но, возможно, вторая волна гибридизации произошла 9 тыс. лет назад. Эти оценки следует считать очень предварительными. Чтобы их улучшить, потребуется анализ существенно большего числа геномов, в том числе представителей разных этнических групп (в частности, центральноафриканских пигмеев мбути, южноафриканских — сан и коса).
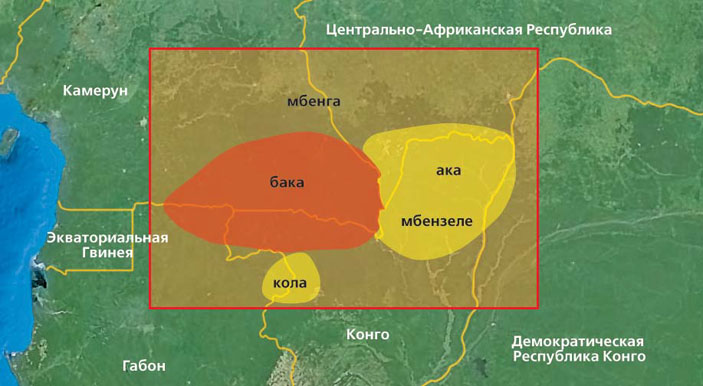
Рис. 4. Области проживания пигмеев (племена бака, ака, мбензеле, кола и мбенга), в геномах которых обнаружились фрагменты, происходящие от неизвестного древнего человека
Наконец, были секвенированы геномы 10 аборигенов Андаманских о-вов (в мировой литературе представленных дикарем Тонга из «Знака четырех» Конан Дойла). Оказалось, что в каждом из них примерно 15 млн нуклеотидов (0,5% размера генома) происходят от неизвестного представителя рода Homo (не от денисовца или неандертальца!), разошедшегося с современным человеком минимум 300 тыс. лет назад [14]. Впрочем, эта работа была подвергнута критике: обнаружились внутренние противоречия между приводимыми статистическими данными и предложенным объяснением, а также отсутствие аналогичного сигнала гибридизации в других наборах данных [15]. В любом случае было бы поучительно сопоставить предполагаемые ультрадревние фрагменты в геномах андаманцев и африканских пигмеев: совпадение даже небольшого числа последовательностей вполне убедительно свидетельствовало бы в пользу их реальности.
Следует отметить, что датировки событий дивергенции и гибридизации, полученные в обсуждаемых работах, слабо согласуются друг с другом, хотя сам факт, что гибридизации неоднократно происходили, уже не подлежит сомнению. Неясно также, до какой степени повышенная доля некроманьонских фрагментов в геномах людей, принадлежащих различным популяциям, вызвана дополнительными гибридизациями, а не другим режимом отбора. Вероятно, с этим можно было бы разобраться, не сопоставляя суммарные статистики, а проводя тщательный дифференциальный анализ геномов гибридизовавшихся неандертальцев и денисовцев, реконструируемых по фрагментам, сохранившимся в современных популяциях, в сравнении со свидетельствами положительного и очищающего отбора в линии кроманьонцев. Впрочем, данных для получения значимых результатов такого анализа сейчас недостаточно — нужны и новые геномы древних людей, и больше разнообразных современных геномов.
Отдельные гены и их группы
Анализ конкретных генов, в отличие от анализа геномов, зачастую приводит к вполне понятным и хорошо согласованным выводам. Пожалуй, самые яркие и многочисленные примеры неандертальского наследства — гены иммунной системы. Кроме уже упоминавшихся в прошлом обзоре гена STAT1 (влияющего на активность врожденного иммунитета), кластера генов OAS (противостоящих вирусным инфекциям) и неандертальских и денисовских аллелей главного комплекса гистосовместимости класса I, была обнаружена высокая частота двух неандертальских и одного денисовского варианта области, содержащей кластер трех генов (TLR6-TLR1-TLR10) Toll-подобных рецепторов, отвечающих за врожденный иммунитет (рис. 5) [16]. Та же область, а также гены IFITM1-3 (кодирующие эффекторные молекулы иммунитета) имеют повышенную частоту неандертальских вариантов у европейцев, а гены NLRC5 (кодирующий Nod-подобный рецептор, участвующий в регуляции врожденного иммунитета), IRF6 (кодирующий регуляторный фактор интерферона) и SIRT1 (кодирующий белок сиртуин-1) — у азиатов, причем в случае TLR6-TLR1-TLR10 и SIRT1 были также найдены независимые признаки положительного отбора — в сторону ускорения эволюции [17].
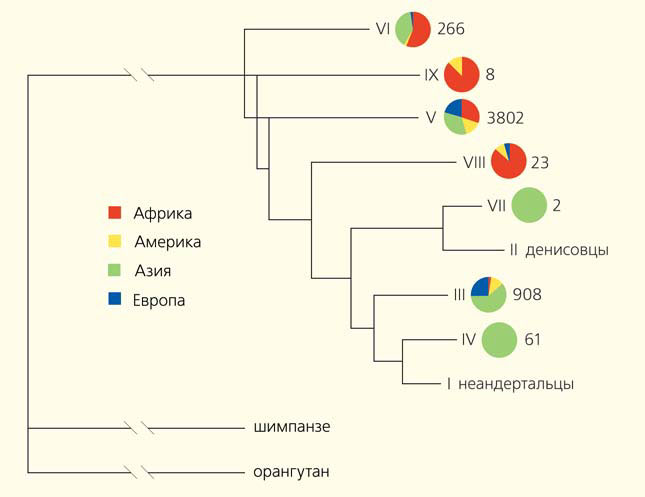
Рис. 5. Филогенетическое дерево вариантов кластера генов TLR6-TLR1-TLR10, кодирующих Toll-подобные рецепторы: семи современных вариантов (III–IX), неандертальского (I) и денисовского (II) [16]. В круговых диаграммах представлено распределение современных вариантов по частям света. Справа от каждой диаграммы обозначено, сколько индивидов имеет такой гаплотип
При систематическом анализе геномов меланезийцев была, в частности, обнаружена повышенная частота неандертальских аллелей генов GBP4 и GBP7, экспрессия которых индуцируется интерфероном, а продукты — гуанилат-связывающие белки — участвуют в реализации механизмов врожденного иммунитета [11].
Систематический анализ вариантов, связанных с генетическими болезнями или предрасположенностями к ним, показал, что с неандертальским наследством преимущественно связаны нейропсихиатрические, кожные и иммунные заболевания [18]. Это неудивительно: коль скоро такие аллели сохранились до сих пор, они, видимо, были адаптивны (или хотя бы нейтральны) в другом контексте. Например, варианты, связанные с нарушениями обмена веществ, могли быть полезны при нерегулярном режиме питания. Особый интерес представляют неандертальские варианты генов, ассоциирующиеся с психическими расстройствами, в частности с депрессией. С одной стороны, известно, что у человека ее развитие связано с нарушением циркадных ритмов, а соответствующие гены перепредставлены в наборе генов, имеющих неандертальские варианты. Однако можно предположить, что эти варианты были адаптивны при переходе от жизни в тропиках к жизни в северных широтах с коротким световым днем зимой. С другой стороны, систематический анализ генов, связанных с шизофренией, показал, что многие из них находились под действием положительного отбора после разделения кроманьонцев и неандертальцев [19].
Итак, как и следовало ожидать, секвенирование новых геномов, как древних, так и современных, усложнило наши представления о предыстории человека. Стало ясно, что гибридизация кроманьонцев с неандертальцами происходила неоднократно, и потомки смешанных пар жили не только с кроманьонцами (став источником неандертальских аллелей у европейцев и азиатов), но и с неандертальцами (объясняя присутствие кроманьонских аллелей у алтайского неандертальца). В геномах некоторых африканцев были найдены следы гибридизации с неизвестным представителем рода Homo. Однако эволюционные сценарии, предложенные в различных статьях, плохо согласуются друг с другом.
Работа выполнена при поддержке Российского научного фонда (проект 14-24-00155).
Литература
1. Гельфанд М. С. Геном неандертальца: открытые вопросы // Природа. 2016. № 1. С. 27–35.
2. Mendez F. L., Poznik G. D., Castellano S. et al. The divergence of Neandertal and modern human Y chromosomes // Am. J. Hum. Genet. 2016. V. 98. P. 728–734. DOI: 10.1016/j.ajhg.2016.02.023.
3. Haldane J. B. S. Sex ratio and unisexual sterility in hybrid animals // Journ. of Gen. 1922. V. 12. P. 101–109. DOI: 10.1007/BF02983075.
4. Wall J. D., Yang M. A., Jay F. et al. Higher levels of neanderthal ancestry in East Asians than in Europeans // Genetics. 2013. V. 194. P. 199–209. DOI: 10.1534/genetics.112.148213.
5. Fu Q., Posth C., Hajdinjak M. et al. The genetic history of Ice Age Europe // Nature. 2016. V. 534. P. 200–205. DOI: 10.1038/nature17993.
6. Rogers R. L. Chromosomal rearrangements as barriers to genetic homogenization between archaic and modern humans // Mol. Biol. Evol. 2015. V. 32. P. 3064–3078. DOI: 10.1093/molbev/msv204.
7. Sawyer S., Renaud G., Viola B. et al. Nuclear and mitochondrial DNA sequences from two Denisovan individuals // Proc. Natl. Acad. Sci. USA. 2015. V. 112. P. 15696–15700. DOI: 10.1073/pnas.1519905112.
8. Meyer M., Arsuaga J. L., de Filippo C. et al. Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins // Nature. 2016. V. 531. P. 504–507. DOI: 10.1038/nature17405.
9. Gibbons A. DNA from Neandertal relative may shake up human family tree // Science. 2015. DOI: 10.1126/science.aad1740.
10. Kuhlwilm M., Gronau I., Hubisz M. J. et al. Ancient gene flow from early modern humans into Eastern Neanderthals // Nature. 2016. V. 530. P. 429–433. DOI: 10.1038/nature16544.
11. Vernot B., Tucci S., Kelso J. et al. Excavating Neandertal and Denisovan DNA from the genomes of Melanesian individuals // Science. 2016. V. 352. P. 235–239. DOI: 10.1126/science.aad9416.
12. Sankararaman S., Mallick S., Patterson N. et al. The combined landscape of Denisovan and Neanderthal ancestry in present-day humans // Curr. Biol. 2016. V. 26. P. 1241–1247. DOI: 10.1016/j.cub.2016.03.037.
13. Hsieh P., Woerner A. E., Wall J. D. et al. Model-based analyses of whole-genome data reveal a complex evolutionary history involving archaic introgression in Central African Pygmies // Genome Res. 2016. V. 26. P. 291–300. DOI: 10.1101/gr.196634.115.
14. Mondal M., Casals F., Xu T. et al. Genomic analysis of Andamanese provides insights into ancient human migration into Asia and adaptation // Nat. Genet. 2016. V. 48. P. 1066–1070. DOI: 10.1038/ng.3621.
15. Skoglund P., Mallick S., Patterson N. et al. No evidence for unknown archaic ancestry in South Asia // 2016. BioRχiv. DOI: 10.1101/068155.
16. Dannemann M., Andrés A. M., Kelso J. Introgression of Neandertal- and Denisovan-like haplotypes contributes to adaptive variation in human Toll-like receptors // Am. J. Hum. Genet. 2016. V. 98. P. 22–33. DOI: 10.1016/j.ajhg.2015.11.015.
17. Deschamps M., Laval G., Fagny M. et al. Genomic signatures of selective pressures and introgression from archaic hominins at human innate immunity genes // Am. J. Hum. Genet. 2016. V. 98. P. 5–21. DOI: 10.1016/j.ajhg.2015.11.014.
18. Simonti C. N., Vernot B., Bastarache L. et al. The phenotypic legacy of admixture between modern humans and Neandertals // Science. 2016. V. 351. P. 737–741. DOI: 10.1126/science.aad2149.
19. Srinivasan S., Bettella F., Mattingsdal M. et al. Genetic markers of human evolution are enriched in schizophrenia // Biol. Psychiatry. 2016. V. 80. P. 284–292. DOI: 10.1016/j.biopsych.2015.10.009.












Рис. 1. Возраст и географическое происхождение образцов ДНК древних кроманьонцев (всего 51) [5]. Каждому геному соответствует столбец, высота которого пропорциональна возрасту найденных останков, а цвет обозначает принадлежность к одному из генетических кластеров. Справа, на краю карты, изображены столбцы для четырех сибирских образцов