Общение микробов
Е. О. Пучков,
доктор биологических наук, Институт биохимии и физиологии микроорганизмов им. Г. К. Скрябина РАН
«Химия и жизнь» №12, 2015
Светлой памяти моего друга Миши Ершова.
Прежде всего следует уточнить, о каких микробах пойдет речь. Дело в том, что микробами или микроорганизмами с давних времен называли все живые существа, видимые только под микроскопом (рис. 1). Постепенно список обнаруженных обитателей микромира расширялся, параллельно шли их изучение и таксономическая классификация. К настоящему времени термин «микробы» объединяет самые разные организмы: бактерии, археи, микроскопические грибы (и их дрожжевая форма, то есть одноклеточная, в отличие, например, от гифальной — нитчатой), простейшие, микроводоросли. Некоторые авторы относят к микробам и микроскопических животных, например коловраток, а также вирусы. Различия между членами этого весьма формального объединения так велики, что каждого из них изучает своя наука: альгология — микроводоросли, протозоология — простейших, зоология — микроскопических животных, вирусология — вирусы, микология — грибы. Бактерии, археи и одноклеточная форма микроскопических грибов — объекты микробиологии. Вот о химической коммуникации у микроорганизмов этих трех групп и пойдет речь.

С легкой руки авторов первого научного обзора в этой области (Kaiser D., Losick R. “How and why bacteria talk to each other” — «Как и почему бактерии разговаривают друг с другом», Cell, 1993, 73, 5, 873–885) коммуникативные химические взаимодействия микроорганизмов исследователи даже в специализированных научных публикациях стали называть «разговорами» или «общением». См., например: Winzer, Hardie, Williams, Current Opinion in Microbiology, 2002, 5, 2, 216–222, “Bacterial cell-to-cell communication: sorry, can't talk now — gone to lunch!” (Общение между бактериальными клетками: извини, сейчас говорить не могу, ушел на обед!); McNab, Lamont, Journal of Medical Microbiology, 2003, 52, 7, 541–545, “Microbial dinner-party conversations: the role of LuxS in interspecies communication” (Разговоры микробов на вечеринке: роль LuxS в коммуникации между видами); Hogan, Eukaryotic Cell, 2006, 5, 4, 613–619, “Talking to themselves: autoregulation and quorum sensing in fungi” (Разговаривая сами с собой: авторегуляция и quorum sensing у грибов).
Многоклеточная жизнь одноклеточных
В естественной среде обитания жизнь микробных клеток протекает в сообществах, которые могут отличаться по плотности популяций: от планктонных форм и небольших конгломератов в водных средах до колоний, биопленок и матов на твердых поверхностях. Еще одна особенность микробных сообществ, как и всего мира микробов, — большое генетическое и метаболическое разнообразие составляющих их видов.
Существует универсальный закон природы, который наиболее кратко сформулировал индейский народ хайда, проживающий на территории Канады и США: «Всё зависит от всего остального». Поэтому естественно ожидать, что в моно- и полимикробных объединениях возникнут межклеточные взаимодействия: физические контакты, химическое взаимовлияние. Однако в силу того же закона химические взаимоотношения микроорганизмов настолько многообразны и так переплетены, что изучение отдельных реакций — непростая задача.
Тем не менее удалось установить, что часть химических взаимодействий имеет определенную специфичность. Так, их можно разделить на антагонистические и коммуникативные. Антагонистические — это, например, секреция антибиотических веществ, подавляющих размножение конкурентных видов. При коммуникативных взаимодействиях секретируемые вещества вызывают модуляцию метаболизма участников, после чего меняются их свойства и/или поведение.
Первыми привлекли внимание исследователей антагонистические отношения между микроорганизмами. Еще Луи Пастер отметил, что это явление, возможно, удастся использовать для борьбы с болезнетворными бактериями. Действительно, были обнаружены и получены для медицинских целей продуцируемые бактериями и грибами антибиотики. Установлено, что некоторые бактерии синтезируют бактериоцины — (поли)пептиды, обладающие антибактериальной активностью с узким спектром действия. Известны так называемые полипептидные киллер-токсины — их выделяют дрожжевые формы некоторых грибов. Несмотря на грозное название, эти вещества специфически поражают только чувствительные клетки того же вида или близкого рода грибов. Противогрибковой активностью обладают также и некоторые секретируемые дрожжевыми грибами гликолипиды.
О существовании коммуникативных межклеточных взаимодействий в мире микробов стало известно сравнительно недавно...
...И было это так
В 1956 году в журнале Nature была опубликована небольшая заметка, в которой сообщалось о взаимовлиянии без непосредственного контакта перед спариванием (англ. mating) двух клеток дрожжей Saccharomyces cerevisiae противоположного типа спаривания (рис. 2; подробно о том, как дрожжи делают это, мы поговорим дальше). Было высказано предположение, что реакцию спаривания регулируют некие «гормоны», секретируемые клетками. Вскоре это заключение получило экспериментальное подтверждение, а индукторы спаривания изолировали в чистом виде. Так были заложены основы обширного научного направления — исследований межклеточной коммуникации при спаривании у разных видов дрожжей. Молекулы, запускающие эти и другие межклеточные реакции, стали называть феромонами, по аналогии с сигнальными веществами многоклеточных животных.

Первые указания на то, что бактериальные клетки специфически воздействуют друг на друга химическими факторами, были получены в 1960-х годах. Стало известно, что стрептококки и бациллы выделяют в среду, в которой их культивируют, соединения пептидной природы, обеспечивающие переход клеток в состояние компетентности для генетической трансформации (такие клетки готовы поглощать ДНК из среды, чтобы приобрести новые, возможно, полезные гены). Позднее некоторые из них были выделены и химически охарактеризованы.
Уникальное явление среди бактерий — жизненный цикл семейства Myxococcaceae, в котором доминирует «социальное» поведение. На стадии вегетативного развития клетки миксококков питаются другими бактериями, перемещаясь и атакуя их большими группами. При голодании на плотной среде клетки начинают координированно двигаться в одном направлении и образуют скопления — так называемые плодовые тела (рис. 3). В плодовых телах часть клеток дифференцируется в сферические споры, устойчивые к нагреванию и высушиванию. В 1962 году была разработана методика, которая позволила смешивать мутантов с различными дефектами формирования плодовых тел и клетки дикого типа. С помощью этой методики удалось установить, что для формирования плодового тела необходимы специальные вещества, секретируемые клетками, а затем идентифицировать пять таких веществ.

В начале 1970-х у люминесцирующих бактерий Photobacterium (ныне Vibrio) fischeri и Beneckea (Vibrio) harveyi (рис. 4) была обнаружена активация биосинтеза люциферазы по мере увеличения концентрации клеток в среде. Исследователи предположили, что бактерии выделяют в среду специфические вещества — автоиндукторы. Вскоре эти вещества были выделены и определена их структура, а само явление назвали quorum sensing, что можно перевести как «чувство кворума». (Далее это выражение будет использоваться без перевода, как термин, подобно латинским in vitro, in situ и т. п.) Когда концентрация клеток достигает определенного уровня («кворума»), концентрация автоиндукторов становится достаточной, чтобы запустить синтез люциферазы и соответственно свечение во всей популяции.


В уже упомянутом обзоре Кайзера и Лозика автоиндукция у люминесцирующих вибрионов рассматривалась наряду с другими проявлениями «коллективного» поведения у бактерий Myxococcusxanthus, Bacillus subtilis, Streptomyces spp., а также у слизевика Dictyostelium discoideum. При этом авторы проводили аналогию с межклеточными сигнальными реакциями у многоклеточных организмов, обеспечивающими регуляцию дифференциации, поведения и морфогенеза.
С тех пор интерес к проблеме межклеточной коммуникации у микроорганизмов, в особенности к quorum sensing, стремительно растет. Исследования развиваются как вширь — увеличивается список микроорганизмов, у которых выявлены коммуникативные отношения, так и вглубь — изучаются их молекулярные механизмы.
Однако результаты практически всех работ по изучению коммуникативных отношений у микробов получены in vitro, на изолированных культурах. Поэтому со временем возникли два вопроса: в какой мере эти реакции проявляются в природных условиях и какие из них следует считать сигнальными?
Сигнализация или не сигнализация
В отношении сигнализации и сигнальных молекул у микроорганизмов сформировалось две позиции, которые условно назовем «либеральной» и «радикальной». Сторонники «либеральных» взглядов считают, что к сигнальным можно отнести любые природные соединения, в том числе микробного происхождения, способные в низких концентрациях вызывать модуляцию транскрипции. В частности, известно, что некоторые антибиотики в низких концентрациях могут модулировать метаболизм микроорганизмов, не подавляя их размножения; в рамках «либеральной» концепции такие антибиотики — сигнальные молекулы. Взаимовлияние бактерий и клеток хозяина-эукариота с участием тех или иных химических факторов также рассматривается как сигнализация между царствами — есть указание на наличие подобных взаимодействий у бактерий с млекопитающими и растениями.
«Радикальная» позиция строится на принципах биологической (эволюционной) целесообразности. Прежде всего ее сторонники предлагают разделить секретируемые коммуникативные вещества на три типа: сигнальные (англ. signal), стимуляторы (cue) и «принудители», или «химические манипуляторы» (coercion). Важным отличием сигнальных молекул считается то, что они возникли в ходе совместной эволюции передающих и принимающих клеток. Сигнальные молекулы переносят информацию, которая вызывает изменения в клетках-реципиентах, благоприятные для развития обоих партнеров. Кроме того, у них выделяют следующие отличительные признаки: 1) ответ, который они вызывают, отличается от физиологических сдвигов, необходимых для метаболизирования или детоксикации данных молекул; 2) их распознают специфические рецепторы клеток-реципиентов; 3) они синтезируются на определенных стадиях развития, при определенных физиологических условиях, и их накопление вызывает согласованный отклик во всех клетках популяции в виде изменения метаболизма после достижения пороговой концентрации.
Есть два типа коммуникативных реакций — индукция спаривания у аскомицетных дрожжей и quorum sensing у бактерий, которые считают сигнальными сторонники обеих позиций, поэтому их иногда называют «каноническими».
Спаривание у дрожжей
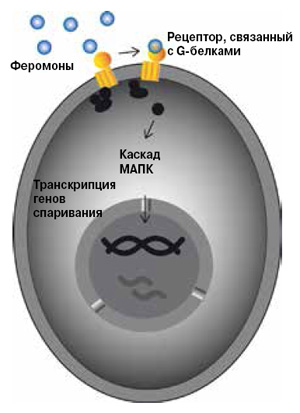
Это явление лучше всего изучено у аскомицетных дрожжей Saccharomyces cerevisiae и Schizosaccharomyces pombe прежде всего потому, что оба вида давно и успешно служат моделями клеток высших эукариот. Спаривание у дрожжей сопровождается мейозом: клетки из диплоидных превращаются в гаплоидные, а при слиянии их восстанавливается диплоидность. Очевидно, в этом и состоит биологическая целесообразность полового процесса у дрожжей — он обеспечивает комбинационную изменчивость вида. Хотя дрожжи S. cerevisiae и S. pombe сравнительно давно разошлись в ходе эволюции и заметно различаются по всем основным биологическим свойствам, принципиальная схема механизмов, обеспечивающих спаривание, у них имеет много общего (рис. 5 и 6).
Гаплоидные клетки противоположного типа спаривания (МАТа и МАТα у S. cerevisiae, Р и М у S. pombe) секретируют сигнальные пептидные молекулы — феромоны (факторы а и α у S. cerevisiae, Р и М у S. pombe), которые взаимодействуют с рецепторами на поверхности партнеров. Это активирует связанные с рецепторами G-белки, которые, в свою очередь, способствуют активации фермента митоген-активируемой протеинкиназы (МАПК). С этого момента начинается внутриклеточная передача сигнала: протеинкиназа фосфорилирует белки-мессенджеры — присоединяет к ним фосфатные группы. Они становятся факторами транскрипции, то есть белками, которые регулируют активность других генов, в данном случае многочисленных генов спаривания. В результате клеточный цикл останавливается, индуцируется образование шму, клетки-партнеры сливаются, затем сливаются их ядра, формируется диплоидная зигота. После этого у S. cerevisiae образуются либо диплоидные клетки в результате митоза, либо (в условиях голодания по азоту) гаплоидные споры за счет мейоза. У S. pombe мейотическое деление зиготы приводит к образованию гаплоидных спор.

Молекулярные детали этого сложнейшего механизма уже сравнительно хорошо изучены. Познакомиться с ними, а также с нерешенными вопросами можно в научном обзоре, ссылка на который дана в подписи к рис. 6.
Quorum sensing
Межклеточная сигнализация по механизму quorum sensing к настоящему времени описана у многих видов бактерий и некоторых дрожжевых форм грибов. Есть данные, указывающие на то, что она существует у архей.
Биологическая целесообразность quorum sensing в большинстве случаев предположительно заключается в следующем. Учитывая энергетические затраты, микроорганизмам имеет смысл включать некоторые направления метаболизма только тогда, когда результат может быть достигнут при условии коллективных однонаправленных действий группы клеток. Установлено, что к таким физиологическим реакциям относятся генетическая трансформация у стрептококков, энтерококков и бацилл, биолюминесценция у люминесцирующих вибрионов, спорогенез у бацилл, образование биопленок и биосинтез антибиотиков у бактерий, образование плодовых тел и спорогенез у миксококков, экспрессия факторов вирулентности у бактерий и дрожжевых форм грибов. Всем этим не стоит заниматься в одиночку: результат окупит затраты энергии только в том случае, если много клеток будут делать то же самое. Свечение небольшого количества вибрионов в тканях кальмара слишком слабо, чтобы его заметили, концентрация антибиотика в среде слишком низкая, чтобы подавить размножение конкурента, если антибиотик выделяет мало клеток, и т. п.
Для принятия решения — запускать или не запускать такие энергоемкие процессы — клетки зондируют окружающую среду, синтезируя и выпуская туда сигнальные молекулы. В разных системах их называют автоиндукторами либо феромонами. Когда их концентрация достигает определенного уровня — что возможно либо при увеличении концентрации клеток, либо при наличии некоего диффузионного барьера (например, когда бактерии заключены в эндосомах клеток хозяина), либо и того и другого, — вся популяция клеток воспринимает это как сигнал для специфических изменений экспрессии генов.
У бактерий существует два наиболее распространенных типа сигнальных молекул quorum sensing. У грамотрицательных видов (а) их чаще называют автоиндукторами, и они представляют собой различные N-ацилпроизводные гомосеринлактона (АГЛ), которые синтезируются синтазой LuxI-типа. АГЛ способны проникать через цитоплазматические мембраны, поэтому синтезированные внутри клеток молекулы выходят в среду, накапливаются и возвращаются по градиенту концентрации в клетки. Там они связываются с цитоплазматическими белками LuxR-типа — регуляторами транскрипции quorum sensing генов, обеспечивающих синтез автоиндукторов и молекулярных компонентов для физиологического ответа.
Второй тип сигнальных молекул выявлен у грамположительных бактерий (б) — короткие посттрансляционно модифицированные пептиды (автоиндукторные пептиды, АИП). Их выводят из клеток специальные транспортные системы, локализованные в цитоплазматических мембранах. Сигнал они передают двумя путями. Первый — через гистидинкиназу на мембране. Связывание АИП с рецепторным компонентом фермента сопровождается его автофосфорилированием, после чего фосфатная группа передается непосредственно или через посредников (в зависимости от вида бактерий) фактору транскрипции, который и модулирует соответствующие гены quorum sensing. Второй путь обеспечивается системой транспорта пептидов, через которую АИП могут возвращаться в клетки, связываться с факторами транскрипции и модулировать активность quorum sensing генов, ответственных за образование АИП и осуществление физиологического отклика. Однако, как любят выражаться специалисты в этой области, химический лексикон межклеточного общения у микроорганизмов не ограничивается только этими двумя «словами» (типами молекул). Известны и другие вещества, которые используют микроорганизмы для внутривидовой межклеточной сигнализации.
Особую роль в изучении quorum sensing сыграли обнаружение и характеристика трех систем quorum sensing у морских люминесцирующих грамотрицательных бактерий Vibrio harveyi (в). Здесь все оказалось необычно. Во-первых, удивителен сам факт существования в клетках одного вида сразу трех систем quorum sensing, которые могут функционировать одновременно и контролировать работу генов, ответственных не только за биолюминесценцию, но и за другие процессы: секрецию III типа, синтез металлопротеаз и сидерофоров. Во-вторых, молекулярная организация одной из систем представляет собой своего рода комбинацию систем грамположительных и грамотрицательных бактерий. Автоиндуктор в ней — АГЛ (N-(3-гидроксибутаноил)гомосеринлактон) синтезируемый ферментом LuxM, а его первичный сенсор — рецептор с киназной активностью LuxN. Его называют автоиндуктором первого типа, АИ-1. В-третьих, автоиндуктор другой системы (CAI-1), 3-гидрокситридекан-4-он, а также компоненты его синтеза и рецепции (не показан на рисунке) обнаружились у патогенного для человека Vibrio cholerae и некоторых морских бактерий. Но главный герой в этой истории — система с автоиндуктором второго типа (АИ-2), S-2-метил-2,3,3,4-тетрагидрокситетрагидрофуранборатом. Это продукт синтеза с участием фермента LuxS, а рецептором для него служит LuxPQ, бинарный компонент с киназной активностью. Ген LuxS и АИ-2 не видоспецифичны — они найдены у многих бактерий, и есть гипотеза, что они используются для межвидового общения.


Молекулярные механизмы quorum sensing изучены достаточно подробно (см. врезку). Для дальнейшего рассказа нам важен в первую очередь так называемый автоиндуктор второго типа — АИ-2. Все дело в том, что ген luxS, кодирующий фермент LuxS, который отвечает за синтез АИ-2, и само это вещество были обнаружены у многих бактерий разных таксономических групп, как грамположительных, так и грамотрицательных. Это породило предположение о том, что автоиндуктор АИ-2 — универсальная молекула межвидовой сигнализации, в терминах «межклеточного общения» — своего рода интернациональное слово, существующее во всех языках. Так родилась...
Гипотеза «своих» и «чужих»
Согласно этой гипотезе, у бактерий может быть два варианта межклеточной сигнализации в формате quorum sensing. Первую осуществляют высокоспецифичные системы, использующие в качестве сигнальных молекул АГЛ и АИП, — это внутривидовая сигнализация, которая позволяет клеткам контролировать концентрацию «своих». Второй вариант, основанный на производстве и детекции АИ-2, служит для межвидовой сигнализации — с его помощью клетки оценивают наличие и концентрацию «чужих» в своем окружении. Гипотеза послужила стимулом к поиску и изучению межвидовых взаимоотношений микробов в различных сообществах. Описаны многочисленные примеры, скажем осторожно, взаимовлияния через различных химических посредников между бактериями разных видов, а также между бактериями, грибами, растениями и млекопитающими.
Следует отметить, что эта гипотеза не раз подвергалась критическому рассмотрению, особенно в той части, которая касается межвидовой сигнализации с участием АИ-2. По мнению сторонников «радикального» представления, имеющихся экспериментальных данных недостаточно, чтобы считать АИ-2 сигнальной молекулой. Фермент LuxS, синтезирующий АИ-2, действительно найден у разных бактерий, но проблема в том, что он участвует в метаболическом цикле активированного метила. Поэтому в экспериментах по изучению роли этого фермента в межвидовом взаимовлиянии трудно отличить сигнальную реакцию, связанную с модуляцией транскрипции, от прямого влияния на метаболизм. Открытым в большинстве случаев остается и вопрос о том, в какой мере взаимодействие клеток разных видов с участием АИ-2 выгодно обоим и может ли оно быть результатом их совместной эволюции. Учитывая эти и некоторые другие аргументы, молекулы, ответственные за кооперативное межвидовое взаимодействие бактерий, пока предлагается относить к категории стимуляторов (cue), а не сигнальных.
Дебаты относительно роли системы LuxS/AИ-2 в межклеточной коммуникации бактерий и точного термина, который следует к ней применять, весьма важны. Исследований химической природы «социального» поведения микроорганизмов все больше, и необходим критический взгляд на них. И конечно же нужно договориться о терминах, чтобы не называть принципиально разные явления одним и тем же словом.
Заключение
Изучение межклеточной коммуникации в мире микроорганизмов — быстро развивающаяся многоплановая междисциплинарная область. Мы лишь коротко представили основные черты ее наиболее крупных направлений. Есть и другие направления, пролегающие рядом с «магистральными путями», что, однако, не умаляет ни их достоинств, ни значимости. Это исследования так называемых бактериальных цитокинов, или, в другой терминологии, факторов, ускоряющих оживление (англ. resuscitation-promoting factors), низкомолекулярных ауторегуляторных факторов, а также регуляции аммиаком развития клеток дрожжей в колониях.
Особую значимость исследованиям сигнальных межклеточных отношений у микроорганизмов придает то, что они создали новый базис для решения многих фундаментальных и прикладных проблем. Наметилась тенденция смены одной из основных парадигм микробиологии, согласно которой жизнеспособность каждой микробной клетки обеспечивается исключительно ее индивидуальными свойствами. Все очевиднее существенная, а в ряде случаев определяющая роль в жизни микробов кооперативного, «социального» поведения. Есть указания на то, что феномен «некультивируемости» в лаборатории многих микроорганизмов из природных образцов может быть связан с межвидовой сигнализацией. Изучается также эволюционное значение quorum sensing: возможно, это был один из первых шагов в появлении многоклеточных. Предлагается использовать quorum sensing у бактерий в качестве модели для изучения эволюционных аспектов социального поведения в биологии. Большие надежды связывают с разработкой новых антимикробных средств на основе ингибиторов систем quorum sensing, ответственных за факторы вирулентности у патогенных для человека и животных бактерий. Это направление особенно актуально из-за стремительно возрастающего числа устойчивых к антибиотикам патогенов. В биотехнологии уже нашли применение основанные на компонентах quorum sensing системы для производства биохимических препаратов, создания искусственных тканей и биосенсоров, ферментации с использованием смешанных микробных культур. А специалисты, работающие в этой области, единодушно признают, что главные практические результаты изучения межклеточного общения микробов еще впереди.










