Эволюционные метаморфозы туникат
Ольга Иванова-Казас
«Природа» №5, 2014
 Об автореОльга Михайловна Иванова-Казас — доктор биологических наук, лауреат премии имени А. О. Ковалевского АН СССР (1958), премии за лучшую публикацию в журналах, издаваемых Международной академической издательской компанией «Наука / Интерпериодика», и медали А. О. Ковалевского Санкт-Петербургского общества естествоиспытателей (2001). Основные научные интересы связаны со сравнительной и эволюционной эмбриологией беспозвоночных животных. Автор нашего журнала с 1936 г. |

Как известно, в мифологии содержится очень много чудесных превращений — метаморфозов. Например, Афина Паллада — древнегреческая богиня мудрости и женских рукоделий — однажды была оскорблена дерзостью молодой ткачихи Арахны, которая осмелилась вызвать ее на соревнование. После этого Арахна соткала холст, на котором были изображены любовные похождения богов и некоторые их неблаговидные поступки. Что выткала сама Афина, неизвестно, но она была так разгневана, что превратила Арахну в паука, который теперь может ткать только паутину. А зоологи стали употреблять термин «метаморфоз» в тех случаях, когда в индивидуальном развитии животных происходят особенно сильные изменения — например, при превращении червеобразной гусеницы в крылатую бабочку. Но еще более значительные метаморфозы произошли в эволюции низших Хордовых* животных — в подтипе Оболочников (Tunicata).
Проблема происхождения и ранней эволюции типа Хордовых начала обсуждаться еще в середине XIX в., причем высказывались подчас совершенно фантастические гипотезы. Разные авторы выводили Хордовых от Немертин, Головоногих моллюсков, Кольчатых червей, Членистоногих, Полухордовых и др.** Заслуживают внимания гипотезы, выводящие тип Хордовых от похожих на головастиков личинок Асцидий путем неотении. Рассмотрим происхождение самой этой личинки.
Асцидий долгое время причисляли к Моллюскам и только после эмбриологических исследований А. О. Ковалевского [1] который изучил строение личинок Асцидий, была установлена их принадлежность к типу Хордовых. Но В. Гарстанг полагал, что «головастик» Асцидий образовался в результате модификации диплеврулы — ранней стадии билатерально-симметричной личинки, присущей Иглокожим и Полухордовым [2]. Но это была чисто умозрительная гипотеза, не подкрепленная никакими фактами [3].

Рис. 1. Схемы строения Ланцетника (а) и Асцидии — на стадии личинки (б) и в конце метаморфоза (в). 1 — анус, 2 — глотка, 3 — жаберная щель, 4 — кишка, 5 — клоакальный сифон, 6 — нервный ганглий, 7 — нервная трубка, 8 — остатки хорды, 9 — прикрепительные сосочки, 10 — ротовой сифон, 11 — сердце, 12 — туника, 13 — хорда, 14 — эндостиль, 15 — эпикард. Мускулатура не изображена
По мнению Н. Дж. Берилла, сначала у Асцидий такой личинки не было [4]. Она возникла внезапно в результате своеобразной мутации, из-за которой клетки дорсальной части энтодермы сильно разбухли, вакуолизировались, расположились одним рядом и превратились в хорду. Хорда стала выпячивать эктодерму, так что образовался хвост, в который были втянуты мышечные элементы и нервная трубка. Но такой сложный орган не мог возникнуть в результате одной-единственной мутации и сразу начать выполнять свою важную биологическую функцию. В настоящее время можно считать общепризнанным, что Асцидии произошли от свободноплавающих Хордовых, но подверглись деградации из-за сидячего образа жизни [3, 5], а самым примитивным среди нынеживущих Хордовых, по-видимому, является Ланцетник — представитель подтипа Бесчерепных, или Головохордовых (Acrania s. Cephalochordata).
Тело Ланцетника имеет удлиненную сплющенную с боков форму (рис. 1, а). По оси его тела проходит упругая хорда, над ней — нервная трубка, а по бокам от нее располагаются продольные мышцы, благодаря сокращениям которых тело изгибается из стороны в сторону. Под хордой лежит кишечник с ротовым отверстием на переднем конце и анальным близ заднего конца. Передняя часть кишки (глотка) расширена, в ее стенках имеются жаберные щели, ведущие в околожаберную (перибранхиальную) полость, которая открывается наружу отверстием на брюшной стороне тела. Ланцетник питается, профильтровывая воду через жаберный аппарат. Во время эмбрионального развития у Ланцетника образуется три пары целомических полостей (прото-, мезо- и метацели); позднее в результате поперечного деления метацелей образуется еще много так называемых постларвальных сегментов.
Асцидии
От животных, близких к ланцетнику, произошел класс Ascidiacea, входящий в подтип Tunicata, который назван так потому, что кожный эпителий этих животных выделяет на своей поверхности студенистый или более плотный слой — тунику, которая выполняет защитную и опорную функцию и позволяет нежным животным сохранять свою форму. В тунику часто внедряются цилиндрические или разветвленные выпячивания стенки тела (сосуды туники) и проникают мезенхимные клетки. Возникновение туники потребовалось и стало возможным после перехода к седентарности, а при плавании она была бы только помехой. А новое название подтипа Urochordata нельзя признать удачным, так как у Сальп и Пиросомид нет ни хвоста, ни хорды.
Тело Асцидий имеет мешкообразную форму, ротовой и клоакальный сифоны занимают на нем апикальное положение (см. рис. 1, в). Хорды и нервной трубки у взрослых асцидий нет, имеется только нервный ганглий, расположенный подле ротового сифона. Кишечник петлеобразно изогнут, его задний конец открывается в перибранхиальную полость, которая играет роль клоаки (в нее впадают и протоки половых желез). В первичной полости тела находится пульсирующий пузырек (сердце), который считается дериватом протоцеля; мезоцелям соответствуют эпикарды (одно или два выпячивания стенки глотки), играющие важную роль при некоторых формах бесполого размножения, а метацелей у взрослых асцидий нет. А так как при сидячем образе жизни встреча особей противоположного пола затруднена, Асцидии стали гермафродитами, и у многих из них возникло бесполое размножение и колониальность. Эмбриональное развитие Асцидий имеет «установку» [6] на стадию личинки, почему удобнее сначала рассмотреть строение этой стадии.

Колониальная асцидия Nephtheis fascicularis. Большой барьерный риф
Выходящая из яйцевых оболочек ювенильная стадия приобрела у Асцидий статус личинки, выполняющей две важные функции — расселения и выбора подходящего места для прикрепления. В ее организации сохраняются все основные черты плана строения Хордовых, но в то же время появились признаки, связанные с ее функцией и подготавливающие предстоящий метаморфоз. Тело личинки разделено на два отдела — на переднее расширенное туловище, в котором сосредоточены зачатки дефинитивных органов, и узкий хвост, выполняющий локомоторную функцию (см. рис. 1, б). В хвосте содержатся хорда и нервная трубка, передний конец которой, находящийся в туловище, расширяется и содержит органы чувств, которых еще не было у Ланцетника, — глаз и статолит (по-видимому, это исключительно личиночные органы). А по бокам от хорды располагаются две мышечные ленты, состоящие из немногих крупных клеток. В области хвоста туника образует спинной и брюшной плавники. На переднем конце личинки находятся органы прикрепления — три чувствительных сосочка или присоски. Из-за них ротовой сифон сдвинут на спинную сторону, соответствующим образом повернут и весь комплекс внутренних зачатков. Эти зачатки у личинок разных видов достигают разного уровня развития, часто, но не всегда уже имеются жаберные щели. Перибранхиальная полость развивается из двух впячиваний эктодермы, которые объединяются на спинной стороне и образуют клоаку, открывающуюся наружу сифоном. Часто у личинок уже имеется сердце и эпикард, а стенки метацелей превратились, по-видимому, в мускулатуру хвоста.
Особенно продвинутыми в своем развитии бывают некоторые живородящие колониальные Асцидии, личинки которых нередко уже приступают к бесполому размножению (рис. 2, а), но сами они еще не питаются, их сифоны остаются закупоренными туникой. Активно плавают они очень недолго — всего один-два дня или несколько часов, после чего прикрепляются к субстрату и претерпевают метаморфоз, в ходе которого хвост втягивается, а содержащиеся в нем органы подвергаются гистолизу.
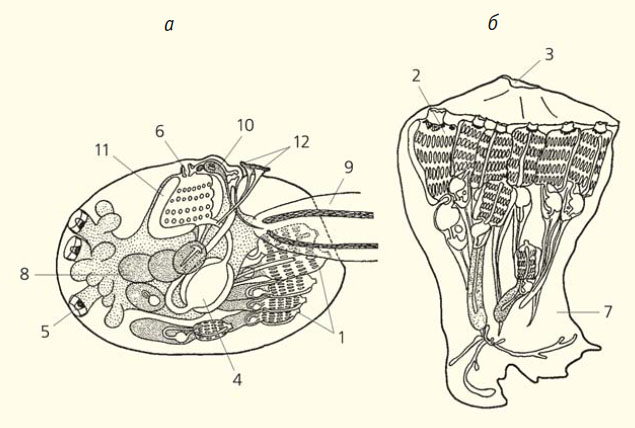
Рис. 2. Асцидия Hypsistozoa fasmeriana: а — личинка; б — молодая колония [7]. 1 — бластозооиды, 2 — оозооид, 3 — общее клоакальное отверстие, 4 — кишечник оозооида, 5 — прикрепительные сосочки, 6 — ротовой сифон, 7 — стебелек, 8 — фрагменты, отделяющиеся от столона, 9 — хвост, 10 — чувствительный пузырек, 11 — эндостиль, 12 — энтодермальные трубки (часть плаценты)
Ранние стадии эмбрионального развития у Асцидий характеризуются строгой детерминацией. Кроме того, эмбриональное развитие сильно ускорено, количество митотических циклов, предшествующих гаструляции, сокращено. У Ланцетника гаструляция начинается, когда зародыш состоит приблизительно из 1000 клеток, а у Асцидий — на стадии 128 бластомеров, т. е. на три-четыре митотических цикла раньше. В зачатках хорды и мышц деление клеток рано прекращается. Количество клеток в мышечных лентах у разных видов довольно сильно варьирует, у Dendrodoa grossularia оно сократилось до 80, а хорда у всех личинок состоит из 40 клеток.
После прикрепления личинки находящиеся на ее переднем конце сосочки распластываются по субстрату, оба ее сифона занимают апикальное положение, а весь комплекс зачатков дефинитивных органов тоже поворачивается приблизительно на 45°. Таким образом, характерная для типичных Хордовых передне-задняя ось сменяется у Асцидий новой апико-базальной осью.

Колониальная асцидия Didemnum molle. Большой барьерный риф
Но эволюция жизненного цикла у Асцидий на этом не заканчивается. После метаморфоза у них появляется высокая регулятивная способность, что создает благоприятную почву для возникновения бесполого размножения, которое представлено у них пятью различными формами. Особенно сложную форму приобрело столониальное почкование. Столон — это цилиндрическое выпячивание кожного эпителия на брюшной стороне тела, в которое входит эпикард, а иногда также отростки от других внутренних органов. От наружного конца столона отделяются фрагменты, из которых развиваются новые индивиды. При этом часто возникают сложные жизненные циклы с чередованием нескольких поколений. Особей, осуществляющих половое размножение, называют гонозооидами, развившихся из яиц — оозооидами, а развившихся в результате бесполого размножения — бластозооидами. У большинства колониальных Асцидий все зооиды способны размножаться и половым, и бесполым путем, но существуют и более сложные циклы. Вообще же эволюция жизненных циклов у Туникат представляет интересную главу сравнительной эмбриологии.
Иногда почкование начинается еще у личинки. В этом отношении особенно интересна личинка Hypsistozoa fasmeriana. У этой Асцидии наблюдается плацентарное живорождение, яйца лишены желтка и имеют в диаметре всего 25 мкм, эмбриональное развитие продолжается больше пяти месяцев, а зрелая личика (вместе с хвостом) вырастает в длину до ~7 мм. От брюшной стороны тела этой личинки отходит пролиферирующий столон, который остается лежать в толще туники и подразделяется перетяжками на несколько фрагментов. Таким образом, уже на стадии личинки различается оозооид и несколько бластозооидов, находящихся на разных стадиях развития (рис. 2, а), а после прикрепления к субстрату сразу получается небольшая колония (рис. 2, б) [7].
Нам пришлось уделить так много внимания Асцидиям потому, что они занимают место у самого корня филогенетического древа Туникат. Хотя это группа, бесспорно, процветающая, но у них, как и у других прикрепленных животных, способность к дальнейшей прогрессивной эволюции (т. е. к ароморфозам, по терминологии А. Н. Северцова [8]) утрачена. Но благодаря высокой регулятивной способности от них (путем идиоадаптаций) произошли остальные классы Оболочников, которые вторично вернулись к свободноплавающему образу жизни.
Аппендикулярии

Рис. 3. Строение Аппендикулярии Oikopleura dioica (а) и ее домика (б) [15]. 1 — анус, 2 — буккальная железа, 3 — гонады, 4 — глотка, 5 — желудок, 6 — жаберная трубка, 7 — каудальный ганглий, 8 — ловчая сеть, 9 — нервный тяж, 10 — отверстие, через которое животное покидает домик, 11 — ротовое отверстие, 12 — хорда, 13 — хвост, 14 — церебральный ганглий, 15 — эндостиль. Черными стрелками показано направление тока воды
Аппендикулярии (класс Appendicularia) — это очень маленькие животные, которые живут недолго и почти не растут. По своему строению они очень похожи на личинок Асцидий. Тело их тоже делится на объемистый туловищный отдел (цефалэнтерон) и хвост, основание которого сдвинуто на брюшную сторону, причем продольные оси туловища и хвоста образуют приблизительно прямой угол (рис. 3, а). Строение внутренних органов, по сравнению с таковым у Асцидий, заметно упрощено. Ротовое отверстие находится у Аппендикулярий на переднем конце тела, перибранхиальная полость и клоакальный сифон отсутствуют, а задняя кишка открывается наружу на брюшной стороне. Жаберным щелям у Аппендикулярий соответствуют две боковые жаберные трубки, которыми глотка открывается наружу. Нервная система состоит из церебрального ганглия, содержащего статолит, и нервного тяжа, который тянется по спинной стороне через туловище и хвост. У основания хвоста находится еще одно скопление нервных клеток. Почти все Аппендикулярии гермафродиты, половые железы располагаются в задней части туловища. Хвост имеет такое же строение, как у Асцидий, но лишен хвостового плавника.
Самая своеобразная особенность этих животных заключается в строении туники. Последняя выделяется только в туловищном отделе, она отслаивается и образует просторный домик овальной формы. Само животное помещается у одного конца этого домика (переднего при плавании), но повернуто своим передним концом к противоположному концу домика (рис. 3, б). В передней стенке домика находятся два решетчатых отверстия, через которые вода входит в его полость, а на противоположном (заднем) его конце имеется отверстие, через которое вода выходит. Ток воды, создаваемый колебаниями хвоста и проходящий сквозь домик, приводит к реактивному движению домика вместе с содержащимся в нем животным. А с передним концом самой Аппендикулярии связана сложная сеть, состоящая из нитевидных тяжей туники. Сквозь эту сеть проходит циркулирующая в домике вода (направление ее движения показано стрелками на рис. 3, б), а содержащиеся в ней пищевые частицы отфильтровываются. Этот домик вместе с фильтрующей сетью периодически сбрасывается, после чего вся эта конструкция формируется заново.
Еще в середине ХХ в. некоторые авторы считали Аппендикулярий самыми примитивными Туникатами [9], но более вероятно, что аппендикулярии произошли путем неотении от личинок Асцидий. В последнее время стали считать Аппедикулярий примитивными и молекулярные филогенетики. Однако этой идее противоречит наличие у них признаков высокой специализации. Во-первых, разделение тела на туловище и хвост (причины которого совершенно ясны, когда речь идет о личинках Асцидий), но у Аппендикулярий оно не могло возникнуть внезапно. Во-вторых, невозможно поверить, что туника появилась у Туникат в такой сложной форме (домик и ловчая сеть), а потом превратилась в простой слой, более или менее равномерно покрывающий все тело. В-третьих, гермафродитизм редко встречается у свободно передвигающихся животных и часто возникает у прикрепленных, поэтому естественно предположить, что Аппендикулярии унаследовали его от Асцидий. И наконец, как уже отмечалось, в развитии Асцидий наблюдается тенденция к раннему началу гаструляции и раннему прекращению деления клеток в личиночных органах, которые тоже состоят из небольшого и постоянного для каждого вида количества клеток. Это еще резче выражено у Аппендикулярий: гаструляция происходит у них на стадии 32 бластомеров, хорда состоит только из 20 клеток, а мышечные ленты — из 10 крупных клеток каждая. При такой далеко зашедшей малоклеточности возврат к нормальному размножению клеток становится невозможным. Из этого следует, что Аппендикулярии могли произойти только от личинок Асцидий, достигших преждевременной зрелости, из развития которых выпали стадии прикрепления и метаморфоза.
Боченочники

Рис. 4. Типы зооидов в жизненном цикле Doliolum: гонозооид (а), оозооид (б), гастрозооид (в), форозооид (г) [16, 17]. 1 — анальное отверстие, 2 — глотка, 3 — желудок, 4 — кишечник, 5 — клоака, 6 — клоакальный сифон, 7 — мышечные ленты, 8 — нервный ганглий, 9 — почки, 10 — ротовой сифон, 11 — сердце, 12 — семенник, 13 — спинной отросток, 14 — столон, 15 — статоцист, 16 — эндостиль, 17 — яичник
Гораздо более значительные изменения жизненного цикла произошли у Боченочников (класс Doliolida); тело этих животных имеет форму бочонка, у которого на месте донышек находятся ротовой и клоакальный сифоны, и который перепоясан лентами кольцевых мышц, похожими на обручи, соединяющие доски у бочек. Протекание воды через тело этих животных обеспечивается сокращениями кольцевых мышц (рис. 4, а). Но Долиолиды характеризуются еще и очень сложным жизненным циклом, в котором фигурируют зооиды нескольких типов, различающиеся по структуре и функции и потому носящие разные специальные названия. Знакомство с ними стоит начать ab ovo (‘от яйца’), т. е. с рассмотрения оозооида, развивающегося из оплодотворенного яйца.
Ранние стадии эмбрионального развития изучены у них не так обстоятельно, как у Асцидий, но известно, что в результате гаструляции обособляются энтодерма, хорда, мезодерма и нервная трубка. К концу эмбрионального развития тело зародыша приобретает сходство с личинкой Асцидий (см. рис. 4, а). В суженном хвостовом отделе содержится хорда, состоящая, как у Асцидий, из 40 клеток, а по бокам от нее лежат мышечные ленты, образованные каждая тремя рядами крупных клеток, уже содержащих миофибриллы, а нервная трубка в хвост не заходит (см. рис. 5, б). Еще оставаясь в яйцевой оболочке, зародыш начинает плавать, потом хвост разрушается, а из переднего расширенного отдела тела развивается оозооид, который вскоре приступает к почкованию. Личинка стала Долиолидам ненужной, после того, как и взрослые особи «научились» плавать.

Рис. 5. Некоторые стадии развития Doliolum: поздний зародыш (а), поперечный разрез хвоста (б), блуждающая почка (в), задний конец оозооида с бластозооидами на спинном отростке (г) [14, 16, 17]. 1 — глоточный тяж, 2 — клоакальный тяж, 3 — клоакальный сифон, 4 — мезодермальный тяж, 5 — мышечные клетки, 6 — нервный ганглий, 7 — ротовой сифон, 8 — спинной отросток, 9 — фороциты, 10 — хорда, 11 — эктодерма, 12 — яйцевая оболочка
Оозооид Долиолид становится родоначальником небольшой колонии, поэтому его называют кормилкой (это производное от латинского слова cormus — ‘стебель’, ‘куст’). Все пространство внутри этого бочонка занято обширными глоткой (жаберной полостью) и клоакой, которые разделены косой перегородкой. В этой перегородке имеется четыре пары жаберных щелей. От глотки начинается пищевод, а задний конец кишечной трубки открывается в клоаку. Дорсальнее глотки расположен нервный ганглий, от которого во все стороны расходятся нервы (но настоящей нервной трубки нет). Один из этих нервов связан с органом равновесия (статоцистом), лежащим слева от глотки. Благодаря сокращениям кольцевых мышц оозооид всасывает воду ротовым сифоном и выталкивает ее через клоакальный сифон; это обеспечивает реактивное плавание ротовым сифоном вперед. Половых желез у оозооида нет, но на брюшной стороне находится пролиферирующий столон, от которого отделяются почки. А на спинной стороне над клоакальным сифоном кожный эпителий образует спинной отросток, тоже имеющий отношение к бесполому размножению.
В состав пролиферирующего столона Долиолид входят плотные клеточные тяжи, отходящие от эпикарда, глотки и перибранхиальных полостей; большое количество мезодермальных клеток и даже клетки полового зачатка, отделяющиеся от столона; небольшие фрагменты свободно лежат в тунике. К каждой почке присоединяется несколько очень крупных амебоидных клеток (фороцитов), которые перетаскивают почки и усаживают их на спинной отросток (см. рис. 5, в). У каждой почки образуется некое подобие ножки, прирастающей к спинному отростку, и возникает нечто похожее на эпителиальную плаценту. Установление плацентарной связи между почкой и породившим ее зооидом — явление очень редкое, но встречающееся у некоторых Асцидий.
Почки располагаются на спинном отростке тремя рядами: медианным и двумя боковыми. Из боковых почек развиваются так называемые гастрозооиды (см. рис. 4, в). Они имеют упрощенное строение: очень широкий ротовой сифон и обширную глотку, причем жаберные щели открываются прямо наружу, так как перибранхиальной полости и клоаки у них нет. Но пищеварительная система у гастрозооидов хорошо развита. Сначала они получают питательные вещества через «плаценту» от оозооида, а потом начинают питаться самостоятельно, а оозооид переходит на их иждивение. Но гастрозооиды не способны ни к половому, и ни к бесполому размножению.
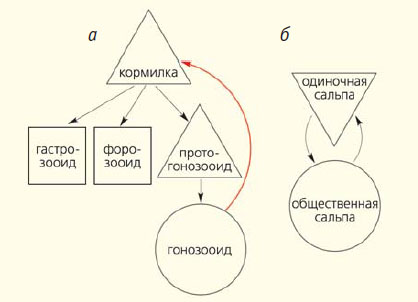
Рис. 6. Жизненные циклы Долиолид (а) и Сальп (б). Название поколения, размножающегося половым путем, помещено в окружности, названия поколений, размножающихся бесполым путем — в треугольниках, а бесплодных поколений — в квадратах. Показан также путь от столона к спинному отростку, по которому перемещаются блуждающие почки
Из медианных почек развиваются форозооиды (см. рис. 4, г). Они имеют бочонковидную форму и первое время остаются прикрепленными к спинному отростку. Форозооиды тоже бесплодны. Почки, отделившиеся от пролиферирующего столона оозооида позднее, прикрепляются уже не к спинному отростку, а к ножкам форозооидов, от которых получают питательные материалы. В этом случае тоже устанавливается плацентарная связь. Зрелые форозооиды отделяются от спинного отростка и начинают плавать и питаться самостоятельно. Из почек, которые они носят на своей ножке, образуются протогонозооиды, которые остаются слабо развитыми, но почкуются и дают начало новому поколению бластозооидов — гонозооидам (см. рис. 4, а). Последние хорошо развиты, имеют около 200 жаберных щелей и половые железы; от них половым путем и происходят оозооиды. Doliolum с бластозооидами на спинном отростке — это, в сущности, полиморфная колония. Сложный жизненный цикл Долиолид изображен в виде схемы на рисунке 6. Как могли возникнуть такие удивительные существа, как Долиолиды? Их происхождение от Асцидий не вызывает сомнений хотя бы уже и потому, что у них имеется эмбрионизированная хвостатая личинка. Конечно, маловероятно, что какая-то взрослая асцидия «вздумала» открепиться от субстрата и начала плавать. По всей вероятности, возникновение класса Doliolida началось с того, что личинка какой-то асцидии перестала прикрепляться, а развитие дефинитивных органов, в отличие от того, что произошло у Аппендикулярий, продолжалось. Это существо приобрело способность самостоятельно питаться и начало расти, при этом хвост оставался по-прежнему маленьким, так как клетки составляющих его органов утратили способность делиться; он уже не мог обеспечить плавание, и предполагаемый предок Долиолид стал передвигаться, с силой выталкивая воду из клоакального сифона. Усовершенствование такого способа передвижения привело к тому, что оба сифона расположились на противоположных концах тела, а превратившийся в бесполезный придаток хвост редуцировался.
Сальпы
По своей организации Сальпы (класс Salpae) очень близки к Долиолидам и оба этих класса часто объединяют под названием Thaliacea (их можно считать сестринскими группами). Главные различия между ними касаются эмбрионального развития и жизненного цикла, который у Сальп (по сравнению с циклом большинства сложных Асцидий) очень прост, в нем чередуются только два, но разных, поколения: оозооида (одиночной сальпы), от которого происходят бластозооиды (общественные сальпы; см. рис. 6, б). Последние размножаются половым путем. У них представлено плацентарное живорождение, при этом каждый бластозооид производит только одно яйцо, а эмбриональное развитие сильно изменилось, и в нем не осталось никаких следов хвоста и хорды [10]. Анатомически оозооид похож на таковой у Долиолид, но у него имеется только одна пара жаберных щелей. В конце эмбрионального развития на брюшной стороне оозооида образуется пролиферирующий столон, в состав которого входят дивертикулы глотки, перибранхиальных полостей, эпикарда, нервной трубки и мезодерма. На поверхности столона появляются перетяжки, подразделяющие его на несколько почек, но эти почки долго остаются связанными друг с другом и отделяются целыми группами, состоящими из 40–65 бластозооидов. Так получаются «общественные» сальпы.
Огнетелки
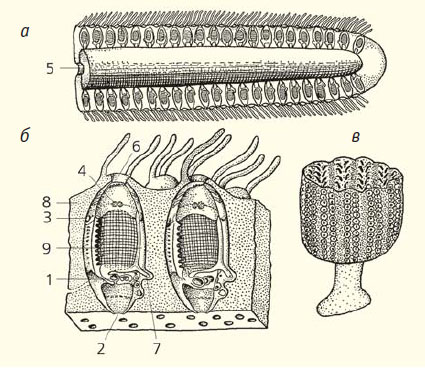
Рис. 7. Колонии Туникат: а — общий вид колонии Pyrosoma; б — часть стенки этой колонии при большем увеличении; в — колония Асцидии Cyathocormus [18]. 1 — кишечник, 2 — клоакальный сифон, 3 — нервный ганглий, 4 — органы свечения, 5 — отверстие полости колонии, 6 — ротовой сифон, 7 — столон, 8 — туника, 9 — эндостиль
Огнетелки (класс Pyrosomida) — существа совсем другого рода (свое название они получили из-за того, что в их теле содержатся светящиеся симбиотические бактерии). Это свободноплавающие колонии, состоящие подчас из многих сотен зооидов, называемых асцидиозооидами. Эти колонии имеют форму полого и открытого на одном конце цилиндра. Стенки этого цилиндра состоят из туники, в них одним рядом располагаются асцидиозооиды (рис. 7, а). По своему строению асцидиозооиды похожи на Асцидий, но сифоны у них обращены в разные стороны: ротовые наружу, а клоакальные — в полость колонии (см. рис. 7, б). Эти колонии тоже плавают реактивным способом — вода всасывается ротовыми сифонами асцидиозооидов, выходит через клоакальные сифоны в общую полость колонии и выводится через ее отверстие наружу. Благодаря этому колония плывет замкнутым концом вперед.
Асцидиозооиды размножаются как половым, так и бесполым путем. Им свойственно яйцеживорождение — крупные богатые желтком яйца развиваются, оставаясь в теле материнского зооида, но не получают от него дополнительного питания. В результате неполного дробления образуется зародышевый диск, из которого формируется оозооид (его называют циатозооидом). Большая часть энтодермы образует зачаток глотки, из выпячивания стенки которой образуется пузырек, обычно называемый перикардом (но не исключено, что это эпикард). А из эктодермы обычным образом развиваются зачатки нервной трубки и двух перибранхиальных полостей. Затем в вентральной части диска начинается развитие пролиферирующего столона, в состав которого входят трубкообразные дивертикулы от всех этих зачатков (рис. 8, 9).
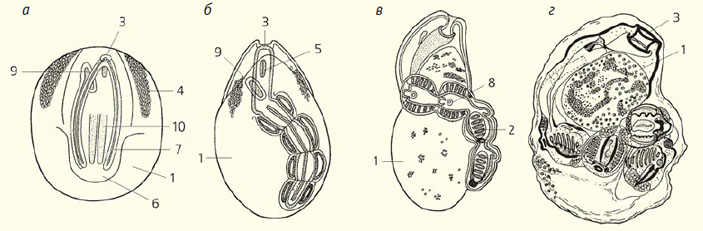

Рис. 9. Поперечный разрез через зародыш Pyrosoma [15]. 1 — вителлофаг, 2 — глоточная трубка, 3 — желток, 4 — мезодермальные клетки, 5 — нервный тяж, 6 — перибранхиальная трубка, 7 — половой зачаток, 8 — эпикард
У большинства изученных Огнетелок столон подразделяется перетяжками на четыре отрезка — будущих основателей колонии. А сам циатозооид остается недоразвитым и постепенно рассасывается вместе с желтком [1, 2]. За счет почкования этих четырех первичных асцидиозооидов начинается формирование колонии, причем сами они располагаются на ее тупом конце.
Иное строение имеет поздний зародыш у Propyrosoma vitiasi, яйца которой содержат очень много желтка и имеют особенно большие размеры (2,1 × 2,5 мм), циатозооид имеет уже строение маленькой асцидии, а столон расчленяется на много фрагментов (иногда — больше 89; рис. 10). Но проследить формирование колонии у этой Пиросомы, к сожалению, не удалось [11].

Рис. 10. Поздний зародыш Propyrosoma: а — внешний вид; б — циатозооид (вид сбоку) [11]. 1 — желток, 2 — жаберные щели, 3 — клоакальный сифон, 4 — нервный ганглий, 5 — пролиферирующий столон, 6 — ротовой сифон
При обсуждении вопроса, каким образом произошли Пиросомы, следует помнить, что в этом случае речь идет об эволюции не отдельных индивидов, а целых колоний. Представить себе превращение цепочки общественных Сальп или полиморфической колонии Долиолид в колонию Pyrosomida очень трудно. А колонии Асцидий обычно стелятся ковром по субстрату, и очень маловероятно, что какая-то из них смогла отделиться от субстрата, свернуться в трубку и начать активно плавать. Но у некоторых Асцидий из семейства Polycitoridae колонии имеют булавовидную форму и сидят на довольно тонкой ножке. Колонии Cyathocormus (см. рис. 7, в) имеют даже форму бокала с обширной внутренней полостью. Особенно важно, что в стенках такого бокала зооиды располагаются так, что их ротовые сифоны обращены наружу, а клоакальные — внутрь, как у Пиросомид. Такой стебельчатой колонии уже не трудно было бы оторваться от субстрата и приспособиться к плавающему образу жизни.

Скопление особей одиночной асцидии Halocynthia aurantium. Японское море
Филогения Туникат
Представляет интерес также вопрос, чем обусловлен возврат четырех классов Туникат к свободноплавающему образу жизни, в чем преимущества последнего? Это можно было бы понять, если бы они стали активными хищниками, но они остаются фильтраторами. И все они, к тому же, используют реактивный способ плавания. Этот вопрос пока остается открытым. Или это, как говорили в старину, «игра природы»?
Все вышеизложенное — результат более чем столетнего изучения Туникат с позиций эволюционной морфологии. Однако в последние десятилетия эти животные заинтересовали также и молекулярных генетиков. Они уже получили интересные результаты, которые, однако, далеко не всегда совпадают с суждениями морфологов. Из обобщающей статьи П. Лемэра [12] мы узнаем, что молекулярная программа индивидуального развития сильно варьирует в зависимости от стратегии размножения туникат, а при бесполом размножении на ранних стадиях эмбриогенеза и бластогенеза у одного вида животных эти программы совершенно различны, они совпадают только после образования основных планов строения.
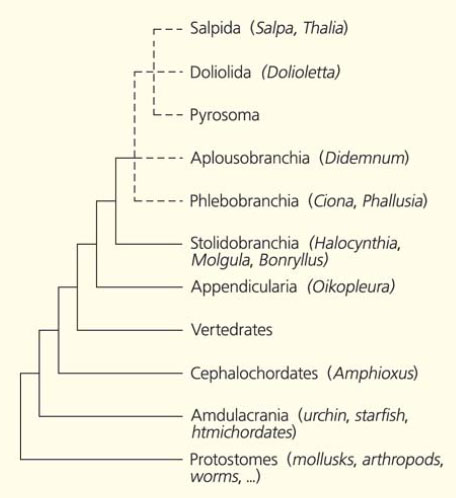
Рис. 11. Кладограмма, изображающая филогенетические отношения между крупными таксонами Deuterostomia [12]
Для выяснения филогенетических отношений между таксонами Туникат используются их молекулярные характеристики. В одной из первых работ на основании анализа последовательностей 18S рРНК был сделан вывод, что Appendicularia — сестринская группа Асцидий из отряда Aplousobranchia, а Thaliacea — из близкого отряда Phlebobranchia [13]. При этом отмечается родство Пиросом и Сальп. Другие авторы на основе анализа того же гена считают, что среди Туникат существует три эволюционных ствола (клады) [14]. К одному из них относятся Aplousobranchia, Phlebobranchia и Thaliacea, ко второму — Асцидии Stolidobranchia, а Appendicularia представляют третий самостоятельный клад.
Филогенетические представления некоторых «молекулярщиков» изображены на рисунке 11. Из этого древа видно, что Аппендикулярии не только самостоятельная, но и самая примитивная группа среди Туникат. Ошибочность этой точки зрения мы уже обсуждали, и возвращаться к этому вопросу не стоит.

Рис. 12. Филогенетические отношения между низшими Хордовыми по данным эволюционной морфологии
Представление о близком родстве Долиолид, Сальп и Пиросомид в общей форме согласуется с данными морфологии; всех их объединяет столониальное почкование, играющее такую важную роль в их жизненных циклах. Но Пиросомид все же следует поставить особняком, так как они, по-видимому, произошли независимо от Thaliacea и на другом конструктивном уровне — на уровне колоний. Таким образом, приведенной кладограмме можно противопоставить схему, составленную на основе данных эволюционной морфологии (рис. 12).
Поскольку развитие всех морфологических признаков начинается с процессов, протекающих на молекулярном уровне, очень важно и интересно было бы знать, как менялась работа молекулярно-генетического аппарата Асцидий в связи с изменениями в их организации и жизненном цикле. Аналогичные вопросы касаются и других классов Туникат — например, что утратили Аппендикулярии вместе с метаморфозом? Ответы на подобные вопросы нужны не только для изучения Туникат, но и для установления взаимопонимания между морфологами и молекулярными биологами.
***
В заключение я позволю себе заметить, что ни один из мифологических метаморфозов по глубине происшедших изменений не может сравниться с преобразованием Хордовых в Сальп или Пиросомид. Кроме того, мифологические метаморфозы часто приводят к образованию совершенно нелепых и нежизнеспособных чудовищ, а эволюционные метаморфозы протекают, хотя и медленно, но под контролем естественного отбора, так что получаются гармонические и жизнеспособные существа.
Литература
1. Kowalevsky A. O. Entwicklungsgeschichte der einfachen Ascidien // Mеm. Acad. Sci. St.-Petersburg. 1866. Ser. 7. T. 10. P. 1–16.
2. Garstang W. The morphology of Tunicata and its bearing on the phylogeny of Chordata // Quart. J. Mikrosc. Sci. 1928. V. 72. H. 1. P. 51–187.
3. Иванова-Казас О. М. Очерки по филогении низших хордовых. СПб., 1995.
4. Berrill N. J. The origin of Vertebrates. Oxford, 1955.
5. Иванова-Казас О. М. Сравнительная эмбриология беспозвоночных животных. Низшие хордовые. М., 1978.
6. Иванов П. П. Общая и сравнительная эмбриология. М., 1937.
7. Brewin D. I. The growth and development of a viviparous compound ascidian, Hypsistozoa fasmeriana // Quart. J. Microsc. Sci. 1956. V. 97. P. 435–454.
8. Северцов А. Н. Морфологические закономерности эволюции. М., 1939.
9. Fenaux R. Les Appendiculares des mers d’Europe et du Bassin Mediterranean. Paris, 1967.
10. Sutton M. F. The sexual development of Salpa fusiformis (Cuvier) // J. Embr. Exp. Morph. 1960. V. 8. P. 268–290.
11. Иванова-Казас О. М. К вопросу об эмбриональном развитии огнетелок (Tunicata, Pуrosomida) // Зоол. журн. 1956. Т. 35. С. 193–202.
12. Lemaire P. Evolutionary crossroads in developmental biology: the tunicates // Development. 2011. V. 138. P. 2143–2152.
13. Stach T., Turbeville J. M. Phylogeny of Tunicata inferred from molecular and morphological characters // Molecular Phylogenetics and Evolution. 2002. V. 25. № 3. P. 408–428.
14. Tsagkogeorgia G., Turon X., Hopcroft R. R. et al. An updated 18S rRNA phylogeny of tunicates based on mixture and secondary structure models // Evolutionary Biology. 2009. V. 9. P. 187–203.
15. Korner W. F. Untersuchungen uber die Gehausebildung bei Appendicularien (Oikopleura dioica) // Z. Morph. Oecol. Tiere. 1952. Bd. 41. H. S. 1–53.
16. Korschelt E., Heider K. Lehrbuch der vergleichenden Entwicklungsgeschichte der wirbellosen Tiere. Jena, 1910. Bd. 2. Abschn. 4. S. 471–896.
17. Neumann G. Cyclomyaria // Handbuch der Zoologie. 1935. Bd. 5. H. 2. S. 324–400.
18. Oka A. On Cyathocormus mirabilis nov. gen., nov. spec., the type of a type of new family of compound ascidian from Japan // J. Coll. Sci. Tokyo. 1913. V. 32. Art. 12. P. 1–30.
19. Godeaux J. Contribution а la connaissance des thaliacés (Pyrosome et Doliolum) // Annals Soc. R. Zool. Belg. 1957. V. 8. P. 1–285.
20. Kowalevsky A. O. Üeber die Entwicklungsgeschichte der Pyrosomen // Arch. micr. Anat. 1875. Bd. 11. S. 597–635.
* По просьбе автора русские названия групп животных приводятся с прописной буквы, а не со строчной, как это принято в настоящее время. — Примеч. ред.
** До сих пор проблемы происхождения и ранней эволюции животных (в том числе хордовых) вызывают жаркие дискуссии у специалистов — классических зоологов и молекулярных генетиков. Подробнее см.: Малахов В. В. Революция в зоологии: новая система билатерий // Природа. 2009. № 3. С. 40–54 (PDF, 5 Мб). — Примеч. ред.












Здесь и далее фото А. А. Семенова