Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Мешают ли хищники друг другу?
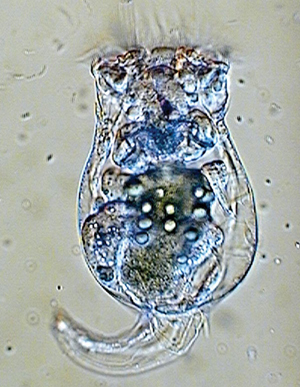
Поедание одних живых существ другими, как то ни печально, является одним из основных типов межпопуляционных взаимоотношений. Скорость потребления хищниками своих жертв (так называемая трофическая функция) зависит как от количества доступных жертв, так и от численности и равномерности распределения в пространстве хищников. Лабораторные эксперименты по питанию коловраток водорослями помогли подобрать значения параметров в трех математических моделях трофической функции и оценить адекватность каждой из них.
Основой всех моделей динамики трофических (пищевых) отношений является понятие трофической функции. Трофическая функция описывает средний рацион хищника – количество жертв, поедаемых одним хищником в единицу времени. Вид функциональной зависимости между рационом хищников и обилием их жертв играет большую роль при изучении механизмов жизнедеятельности экосистемы. В общем речь идет, скорее, о связи ресурсов и потребителей, поскольку не только хищники и жертвы, но паразиты и их хозяева, травоядные животные и их пища – это все примеры взаимодействия типа «хищник-жертва». Выбор той или иной модели функциональной зависимости и соответствующего набора параметров приводит к разным типам временной динамики жертв и организмов, их потребляющих. Соответственно меняются и прогнозы реакции экосистемы на какие-либо воздействия.
Несмотря на то, что первые варианты трофической функции были предложены в 20-х годах ХХ века, эта тема до сих пор является полем для дискуссий в экологическом моделировании.
Авторы рассматриваемой статьи Ю. В. Тютюнов, Л. И. Титова, Ф. А. Сурков и Е. Н. Бакаева сопоставили ряд моделей трофической функции и для трех из них подобрали параметры с помощью лабораторного эксперимента в системе «микроводоросли – коловратки».
В простейшем случае индивидуальный рацион хищника линейно возрастает по мере увеличения количества жертв и является идеализированным представлением случайных столкновений хищников с жертвами в однородно распределенном сообществе при небольших плотностях обеих популяций. Это – зависимость первого типа (зависимость Лотки-Вольтерра).
Однако при увеличении количества жертв наступает момент, когда хищник может убить жертву, но уже не может ее съесть. Существует потолок, насыщение рациона хищника, сверх которого он жертв убивать не станет. Хищника ограничивает время, затрачиваемое на «обработку» одной жертвы: ее поиск, преследование, поимку, удержание, умерщвление, поглощение и, наконец, переваривание. Функциональная зависимость, этот факт учитывающая, называется функциональной зависимостью второго типа (наибольшую известность и распространение получила модель трофической функции Холлинга).
Но и такая модель учитывает не всё. Например, она не рассматривает влияние плотности популяции самого хищника на скорость потребления им жертв. Наиболее простой вид трофической функции, зависящей от плотности хищника, был предложен Р. Ардити и Л.Р. Гинзбургом в 1989г. Неоднородность распределения популяций тоже воздействует на индивидуальный рацион хищника, а в природе чаще всего как раз можно наблюдать активные пространственные перемещения хищников при целенаправленном поиске своих жертв, агрегирование жертв и групповую охоту хищников (см. также: Судьба сообщества зависит от способности особей предвидеть чужие поступки).
В модели, предложенной авторами, как и в большинстве подобных работ, для простоты рассматривается только негативный эффект повышения плотности хищников – конкуренция особей между собой. Положительное влияние кооперации хищников (например, образование стаи для более успешной охоты) здесь не принимается во внимание.

Из ряда обсуждаемых в статье моделей были отобраны три, учитывающие количество хищников: Беддингтона – ДеАнжелиса (1975 г.), Ардити – Гинзбурга (1989 г.) и «Обобщенная зависимость» (GRD), предложенная авторами этой статьи в 2008 году. Эти три модели были апробированы на данных по питанию коловраток (Brachionus calyciflorus, Philodina acuticornis), посаженных в культуру тех или иных водорослей (Chlorella vulgaris, Scenedesmus quadricauda, Synechocystis sp.). Коловраток каждого вида помещали в культуру каждого из видов водорослей. Причем менялась как концентрация водорослей в среде при неизменной концентрации коловраток, так и наоборот.
Эксперименты показали, что с увеличением плотности коловраток происходит уменьшение скорости потребления ими пищи – даже при низких значениях плотности популяций. Авторы статьи считают, что благодаря трофотаксису коловратки концентрировались в местах скопления водорослей и конкурировали между собой.
Наиболее точное приближение трофической функции к экспериментальным данным оказалось у модели, предложенной авторами – Обобщенной зависимости (GRD). Остальные две функции были лишь незначительно хуже и тоже продемонстрировали высокую корреляцию с натурными данными.

Адекватно подобранные трофические функции позволяют не только рассчитать динамику трофических связей между видами, составляющими сообщество, но и предсказывать развитие инвазий видов-вселенцев или распространение болезней; а с помощью биологического контроля – использования «живого против живого», полезных организмов против организмов неполезных – можно сдерживать рост популяций видов вредителей.
Популярные синопсисы