
Эволюция сквозь призму геномики
Эту книгу стоит прочесть каждому интересующемуся современным состоянием науки о происхождении и развитии жизни на Земле. Естественно, со времен Дарвина представления о биологической эволюции значительно изменились. Примерно два десятилетия назад, когда биологи перешли от изучения отдельных признаков и генов к исследованию полных собраний «генетических текстов» — к анализу геномов, появились недоступные ранее возможности для выдвижения и проверки гипотез.
Как известно, первый бактериальный геном был прочтен (секвенирован) в 1995 г., а первый вирусный — почти на 20 лет раньше. С тех пор благодаря бурному развитию технологии секвенирования расшифрованы последовательности нуклеотидов уже тысяч различных организмов (от мелких вирусов и прокариот до крупных геномов высших растений и млекопитающих, включая человека), причем для некоторых видов собрана генетическая информация от множества отдельных особей, что позволяет оценивать внутривидовую изменчивость на уровне ДНК.
Для анализа накопленных массивов данных о структуре геномов, регуляции генов, взаимодействии молекул РНК, белков и метаболитов разработаны изощренные статистические процедуры и методы компьютерного анализа, ставшие основным инструментом новых направлений науки — сравнительной геномики и системной биологии. Полученные результаты надо обобщить. По мнению одного из пионеров этого направления Е. В. Кунина (автора книги), такое обобщение потребует трудов еще двух научных поколений. Но уже сейчас он сделал попытку наметить черты будущей теории, выявить области эволюционной биологии, в которых накопленные данные противоречат сложившимся ранее представлениям, и предложить некоторые предварительные решения.
Значительная часть книги основана на собственных работах автора. Евгений Викторович — выпускник кафедры вирусологии биологического факультета Московского государственного университета им. М. В. Ломоносова. В 1983 г. там же защитил кандидатскую диссертацию под руководством известного российского вирусолога В. И. Агола. До 1991 г. работал в московских НИИ, затем переехал в США и ныне возглавляет группу эволюционной геномики в Национальном центре биотехнологической информации (Бетезда, штат Мэриленд).
«Логика случая» сначала вышла на английском языке [1], на русский книга была переведена группой энтузиастов, согласовавших свою работу в социальной сети «Живой журнал». «Люди по-прежнему делают важные дела не за деньги», — пишет в своей рецензии один из переводчиков, Д. Тулинов и отмечает, что, хотя вряд ли такой способ должен становиться правилом, в отношении «Логики случая» он стал следствием ее незаурядности. Перевод был отредактирован автором книги и снабжен дополнительными комментариями.
Книга задумана как научно-популярная, но получилась гораздо ближе к научной. Однако язык ее столь ясен, что для понимания основных положений книги требуются лишь базовые знания биологии.
В 13 главах последовательно изложены история и современное состояние эволюционной биологии сквозь призму сравнительной геномики. Основная часть книги дополнена двумя приложениями: философским (о природе и целях научных исследований) и космологическим (описывающим инфляционную модель эволюции Вселенной и теорию «мира многих миров»). Оба приложения автор использует как «контекст», в который он пытается вписать концепцию эволюции жизни.
В предисловии и введении Евгений Викторович признается в своем пристрастии к изучению вирусов, с которых он начал свою научную карьеру, и отмечает ключевые изменения концептуальной структуры биологии, побудивших его написать книгу. К ним относятся отход от представлений о древе жизни как центральной метафоре эволюционной биологии (данные геномики показывают, что в эволюции вирусов и прокариот понятие древа неприменимо), и о любом эволюционном событии как адаптивном (адаптивность надо не подразумевать, а доказывать). Еще одна причина — изменение взглядов на соотношение случайности и закономерности в эволюции. Взаимодействие между случайными и упорядоченными процессами — главная тема книги, и роль случайности в эволюции в свете современных данных оказывается намного больше, чем это можно было представить ранее.
Развитие эволюционных концепций
История эволюционного учения изложена в книге столь четко и ясно, что это описание могло бы украсить любой учебник. В первых главах также объясняется, почему необходимы пересмотр и систематизация представлений об эволюции жизни. Автор проводит детальный анализ того, какие положения дарвинизма и более поздних концепций актуальны поныне, а какие следует поместить в музей истории науки. Анализ этот проведен с акцентом на основную тему книги — соотношение роли случайности и закономерности в эволюции, и для каждой концепции указано, какую роль она отводит случайности.
Термин «дарвинизм» Кунин использует только в исторической части, справедливо отмечая, что термин неудачный — никто не говорит «ньютонизм» или «энштейнизм», а например «морганизм» с различными дополнениями можно найти исключительно в идеологизированном негативном контексте. Называть современную эволюционную биологию дарвинизмом — смысловая и историческая ошибка.
Автор книги формулирует три важных обобщения, сделанных Дарвином. В отличие от теорий современников, его естественно-научное объяснение эволюции не привлекало телеологические силы или стремление к совершенствованию. Дарвин предложил конкретный механизм эволюции, основанный на взаимодействии между наследственной изменчивостью и естественным отбором. Идеи эволюции он расширил на всю историю жизни, которая была представлена древом, восходившим к общему корню. В построениях Ламарка случайность отсутствовала, а по Дарвину изменчивость случайна, а отбор направлен и создает сложность. Противоречия, с которыми столкнулась его теория в XIX в., включали оценку возраста Земли (проблема разрешилась в начале ХХ в. — после открытия радиоактивности оценка была увеличена с сотни миллионов до нескольких миллиардов лет). Другая проблема — эволюция сложных органов. Дарвин предположил, что она идет через серию промежуточных стадий, каждая из которых частично выполняет функцию развивающегося сложного органа. Еще одной серьезной проблемой был «кошмар Дженкина», названный так по имени читателя, обратившего внимание на то, что благоприятные признаки «растворяются» от поколения к поколению. Дискретная природа «наследственных задатков», открытая Менделем, не была известна Дарвину, и он предложил откровенно слабую теорию пангенеза. Эта проблема исчезла после переоткрытия законов генетики и последующего развития синтетической теории эволюции. Удивительно, но сначала считалось, что генетика опровергает теорию Дарвина. Причиной тому было представление о мутациях как скачкообразных процессах, тогда как постепенные изменения, градуализм, считались неотъемлемой частью дарвинизма.
Объединение дарвинизма и генетики произошло в 1920–1950-х годах. Основы были заложены трудами основателей популяционной генетики — Р. Фишера, С. Райта и Дж. Б. Холдейна. Они разработали модели, в которых эволюции были подвержены группы скрещивающихся особей — популяции. Фишер показал, что интенсивность отбора зависит от размера популяции (точнее, ее эффективного размера — той части, которая оставляет потомство). Райт внес понятие «дрейф генов». Этот процесс играет важнейшую роль в эволюции малой популяции, в которой за счет дрейфа могут закрепиться нейтральные или даже вредные мутации. Тем самым Райт указал на то, что случайность проявляется не только в возникновении изменений, но и в их закреплении. Как формулирует Кунин, дрейф генов забирает у отбора часть ответственности. Однако открытие Райта было упущено в формулировках «современного синтеза». Этот термин, введенный в 1942 г. Дж. Хаксли, используется преимущественно в США, в европейской литературе принято писать «неодарвинизм», в русскоязычной — «синтетическая теория эволюции» (СТЭ). В СТЭ не придавалось значения и еще одной модели, описанной Дж. Мейнардом-Смитом, — случайному закреплению нейтральных или слабовредных мутаций из-за их сцепления с полезной мутацией.
«Современный синтез» (или формулирование СТЭ) был проведен генетиком Ф. Добржанским, зоологом Э. Майром и палеонтологом Дж Г. Симпсоном. Симпсон реконструировал картину эволюции на основе палеонтологической летописи и описал периоды стазиса в эволюции многих видов, чередующиеся с периодами быстрой смены доминирующих видов (концепция квантовой эволюции). Однако это представление не вошло в СТЭ. Кунин кратко резюмирует основные принципы эволюции, кодифицированные в СТЭ, следующим образом: случайные мутации — единственный источник эволюционно значимых изменений; естественный отбор (наряду со случайной изменчивостью) — основная движущая сила эволюции. Результат — фиксация редких полезных и элиминация вредных изменений; полезные изменения малы (градуализм); эволюционные процессы не меняются по существу на протяжении истории жизни и на разных уровнях эволюционных изменений; древо рассматривается как метафора эволюции жизни, из чего следует существование единого общего предка или нескольких форм-предшественников.
Позже С. Дж. Гулд и Н. Элдридж собрали обширные данные, показывающие, что история многих видов, восстанавливаемая по палеонтологической летописи, соответствует состоянию стазиса, перемежающемуся внезапным в масштабах летописи исчезновением одних видов и замещением новыми видами. Сформулированная концепция прерывистого равновесия противоречила градуализму. Постепенности изменений противостояла и разработанная С. Оно концепция эволюции путем дупликации гена. В конце 1970-х адаптационизм СТЭ был атакован Гулдом и Р. С. Левонтиным, которые призывали не только искать адаптационистские объяснения, но и учитывать случайный дрейф, внутренние ограничения и другие факторы.
Отмечая достоинства СТЭ, Кунин называет ее догматичной и удручающе незаконченной теорией, относя к ее недостаткам бездоказательное распространение механизмов и закономерностей микроэволюции на макроэволюционные процессы, а также отсутствие представлений об эволюции микробов и о происхождении жизни. Соотношение достоинств и недостатков СТЭ можно представить с другими акцентами, но именно перечисленные проблемы удается разрешить (или хотя бы наметить пути решения) с использованием геномных данных.
Переход к геномному подходу в изучении эволюции был заложен открытием двойной спирали ДНК Дж. Уотсоном и Ф. Криком. Как изящно формулирует Кунин, они «вывели из структуры ДНК биологическое воплощение общего принципа цифрового хранения, кодирования и передачи информации». При этом принцип комплементарности, или принцип Чаргаффа (напротив нуклеотида аденина встраивается тимин или его эквивалент урацил, если речь идет о РНК, а напротив гуанина — цитозин), работает и при репликации, и при синтезе любых видов РНК, и при синтезе белка (комплементарность кодона мРНК и антикодона тРНК). Из теории информации следует, что «передача информации абсолютно без ошибок невозможна в принципе», а из законов термодинамики — что «снижение уровня ошибок возможно только за счет затраты энергии». Кунин формулирует принцип подверженной ошибкам репликации, впервые описанный М. Эйгеном, в виде обобщения: «Репликация цифровых носителей информации неизбежно подвержена ошибкам, что влечет за собой эволюцию этих носителей путем естественного отбора и случайного дрейфа генов при условии, что уровень ошибок репликации ниже катастрофического порога, имеющего порядок величин от одной до десяти ошибок на геном за один цикл репликации» (с. 40). Затем автор обсуждает верхний и нижний пороги ошибок репликации, обеспечивающие, с одной стороны, возможность сохранения жизни, с другой — разнообразие, достаточное для эволюционного процесса.
Далее следует перечисление ключевых открытий и подходов, легших в основу эволюционной геномики. Это концепция «молекулярных часов» Л. Полинга и Э. Цукеркандля; нейтральная теория молекулярной эволюции М. Кимуры; развитие различных подходов к сравнению нуклеотидных последовательностей генов и геномов, начало чему было положено при анализе вирусных геномов и позже модифицировано при исследовании геномов бактерий для построения филогенетических деревьев; появление критериев, позволяющих выявить действие отбора, при изучении белок-кодирующих последовательностей нуклеотидов (по соотношению замен нуклеотидов, меняющих кодируемую аминокислоту, и так называемых синонимичных замен, не влияющих на последовательность аминокислот). Автор отмечает, что качественная идея Дарвина, выраженная в абстрактной математической форме Фишером и проверенная экспериментально генетиком Добржанским, получила в геномике вид прямо измеряемого статистического параметра, характеризующего последовательность нуклеотидов.
Огромную роль сыграли открытия конца ХХ в., сделанные К. Вёзе и его коллегами. Они обнаружили консервативность рРНК. Она настолько консервативна, что сходство последовательностей нуклеотидов видно от бактерий до высших организмов. Это позволило построить филогенетическое древо рРНК, отражающее взаимоотношения всех клеточных форм жизни. При этом обнаружилось, что прокариоты разделяются на две группы, названные эубактериями (собственно бактериями) и архебактериями (или археями). Результатом этих работ стало трехдоменное древо жизни, включающее домен эукариот и два домена прокариот — бактерий и архей.
Филогенетический анализ ДНК из митохондрий и хлоропластов подтвердил симбиотическую теорию происхождения эукариот, развиваемую Л. Маргулис. По митохондриальной ДНК они оказались сходными с альфа-протеобактериями, а по хлоропластной — с цианобактериями. Симбиотическое происхождение хлоропластов предполагали русские исследователи еще в начале ХХ в., но окончательное подтверждение оно получило при обнаружении сходства геномов этих клеточных органелл с геномами прокариот.
Существенными для развития эволюционной биологии были концепции С. Райта и К. Уоддингтона. Райт ввел понятие адаптивного ландшафта, отображающего результаты математического анализа отбора в виде «адаптивных пиков» и разделяющих их долин. В большой популяции эволюционная траектория быстро выходит на пик адаптивности (на рисунке показано сплошной линией), а в малой популяции дрейф генов может «уводить» популяцию от адаптивных пиков (пунктирная линия). Идея эпигенетического ландшафта принадлежит Уоддингтону. Он независимо нашел сходную форму представления возможного пути развития эмбриона, представленного в виде шарика на склоне, а глубина каналов соответствует вероятности каждой траектории. Наличие каналов обеспечивает устойчивость развития к возмущениям, вызванным факторами среды или мутациями. Внешнее давление может нарушить канализированное развитие («перебросить» шарик в мелкий канал) и обнаружить скрытую изменчивость.

Кунин отмечает концепцию эгоистичных генов, предложенную в 1976 г. Р. Докинзом. Он пришел к выводу, что естественный отбор может действовать не только на уровне организма в целом, но и на уровне индивидуального гена. Автор «Логики случая» не рассматривает в данном разделе другой полюс действия отбора — группу, или, выражаясь более современным языком, многоуровневый отбор. Концепция группового отбора выпала из основного направления эволюционных исследований [3]. При изучении геномов можно анализировать «эгоистичные гены», но для того чтобы геномика «увидела» групповой отбор, надо предположить, как он может отображаться на уровне нуклеотидных последовательностей. Однако групповой отбор упоминается при описании доклеточных этапов эволюции жизни (глава 11).
Избранные автором книги темы охватывают описание эволюции геномов в целом (глава 3 — «Сравнительная геномика: эволюционирующие геномные ландшафты» и глава 4 — «Геномика, системная биология и универсалии эволюции: эволюция генома как феномен статистической физики); ниспровержение древа жизни как основной репрезентации хода эволюционного процесса (глава 5 — «Сетевая геномика мира прокариот: вертикальные и горизонтальные потоки генов, мобиломы и динамика пангеномов» и глава 6 — «Филогенетический лес и поиск неуловимого древа жизни в век геномики»); происхождение эукариот в результате симбиоза археи и бактерии, с изложением оригинальной концепции развития системы внутренних мембран и ряда других характерных для эукариотической клетки особенностей как защиты от инвазии бактериальных генов (глава 7 — «Происхождение эукариот: эндосимбиоз, удивительная история интронов и исключительная важность единичных событий в эволюции»); анализ истоков биологической сложности (глава 8 — «Неадаптивная нулевая гипотеза эволюции геномов и истоки биологической сложности»); эволюцию эволюционируемости с предложением оригинальной схемы взаимодействия режимов эволюции (глава 9 — «Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции»); систематизацию мира вирусов и их эволюции с позиций геномики (глава 10 — «Мир вирусов и его эволюция»); происхождение жизни (глава 11 — «Последний универсальный общий предок, происхождение клеток и первичный резервуар генов» и глава 12 — «Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы»). В заключительной главе автор обобщает изложенные им представления об эволюции жизни и очерчивает возможный облик постсовременной теории эволюции. «Постсовременная» (в английском варианте — postmodern), подразумевает одновременно и «после СТЭ» (после Modern Synthesis) и «постмодернистское» (в смысле объединения, казалось бы, взаимоисключающих представлений). В предисловии автор предупреждает, что книга — сборник глав по избранным темам и не претендует быть всеохватывающим трудом.

Различные модели происхождения эукариот (в рецензируемой книге см. рис. 7-2, с. 209). На «археозойном» древе (а) таксоны «группы кроны» имеют митохондрии, остальные (археозои) считались ранней ветвью эукариот. На «лучевом» древе (б) отсутствие митохондрий в некоторых группах объясняется их утратой (преобразованием в митохондриеподобные органеллы), а не ранним отделением групп. Лишь три супергруппы (Uniconta, Plantae и Chromalveolata) из пяти включают многолекточные организмы. Хотя животные и растения — наиболее заметные формы жизни, огромное разнообразие эукариот определяется одноклеточными формами
Рассмотрим нетривиальные факты, эмпирически обоснованные выводы и четко отделенные от них гипотезы, приведенные в книге. Остановимся на самых ярких примерах, которые иллюстрируют возможности геномного подхода к изучению эволюции.
Развитие геномики и современные эволюционные концепции
Сравнение геномов показало, что сами гены относительно устойчивы (т. е. остаются сходными у различных организмов на протяжении миллиардов лет эволюции), тогда как генный состав и устройство геномов меняются довольно быстро. Почти во всех геномах клеточных организмов представлено «ядро жизни» примерно из 70 высококонсервативных генов. Их доля в каждом конкретном геноме составляет менее 1–10%. От 10% до 30% приходится на малоконсервативные гены (встречающиеся в узкой группе организмов) и те, которые не найдены в других изученных геномах. Остальное — умеренно консервативные гены примерно из 5000 групп (кластеров). Элементарные события эволюции генов и геномов сходны (и, добавим вслед за высказанным в рецензии М. С. Гельфанда сетовании об упущенной лингвистической параллели, соответствуют тем ошибкам, которые делает переписчик в тексте): замене (нуклеотида в гене или замещению одного гена другим в геноме соответственно), потере (делеции), вставке, рекомбинации, дупликации.
Интересное следствие возможности замещения генов (выполнение неродственным геном одной и той же функции) — отсутствие универсального минимального набора генов, обеспечивающего жизнь клетки. Минимальный набор генов может быть реконструирован при сравнении геномов минимального размера (это геномы паразитических бактерий, содержащие немногие сотни генов). Среди них преобладают гены, кодирующие белки трансляции, транскрипции и репликации. Представлены были и белки транспортной системы, и ферменты. Большинство генов из этого списка действительно необходимы для выживания клетки, как показали эксперименты с нокаутом (инактивацией) генов. Но возможность замещения генов приводит к тому, что у разных организмов минимальный набор функций, необходимых для выживания клетки, выполняется самыми разнообразными генными ансамблями.

Общий размер геномов и число генов у вирусов, прокариот и эукариот (в рецензируемой книге см. рис. 3-2, с. 71). Данные опубликованы на веб-сайте Национального центра биотехнологических исследований (США)
Анализ обширных массивов данных о строении генов и геномов приводит Кунина к выводу, что общие закономерности эволюции геномов и молекулярных фенотипов описываются стохастическими процессами (вот оно, царство случая!), ограниченными необходимостью сохранять жизнеспособность клетки (поддерживать «существующую общую, но неспецифичную, архитектонику генома и устройства клеток»). Это сохранение обеспечивается очищающим отбором (кто не сохранил, тот вымер). Адаптивным процессам отводится роль модулятора этих стохастических закономерностей. Происходят они в значительной мере за счет добавления к существующему новых частей из уже имеющихся «подручных материалов». Эта идея, сформулированная Ф. Жакобом, отлично подтверждается на геномном уровне.
Здесь следует отметить, что понимание эволюции и адаптации теми, кто разглядывает нуклеотидные последовательности, порой значительно отличается от представлений тех, кто судит об эволюции с точки зрения «классических» (наблюдаемых вооруженным различными оптическими приборами глазом) признаков, использовавшихся до «геномного переворота». В поле зрения «классических» биологов вообще не попадают ни синонимичные замены нуклеотидов, ни замены аминокислот, не влияющие на функции белка. А занимающиеся геномикой специалисты «видят» как значимые изменения (не всегда имея молекулярные инструменты для оценки их адаптивности или вредности), так и нейтральные, которых на уровне генома, естественно, заметно больше. Здесь Кунин проводит параллель между эволюционной биологией и статистической физикой, отмечая проявление общих статистических принципов в поведении больших ансамблей слабо взаимодействующих объектов. По крайней мере на молекулярном уровне это так и выглядит. А следствия для макроуровня должны обсуждаться дополнительно.
Невозможно не согласится с автором и в том, что важнейшее достижение эволюции — это появление клетки и что большая часть наиболее эволюционно значимых событий произошла в первые сотни миллионов лет существования жизни на Земле, до появления современного типа клеток. Однако утверждение, что все остальное не так уж важно, — пожалуй, преувеличение. Дом без фундамента невозможен, но живем-то мы в доме, а не в фундаменте. К тому же, если когда-нибудь выяснится, как появились репликаторы на основе нуклеиновых кислот, «самое важное» может сжаться до этого самого появления, так как все остальное — следствие реализации на их основе принципа подверженной ошибкам репликации в условиях ограниченной доступности жизненных ресурсов.
Одним из важных следствий расширения в сторону мира прокариот материала, рассматриваемого в эволюционных исследованиях, стало падение метафоры «древо жизни». Да и у высших эукариот, как оказалось, основная часть генома состоит из остатков вирусных геномов (у млекопитающих — на две трети). Кунин подробно разбирает различные сочетания представлений о сетевых компонентах (которые были сформулированы, когда стало ясно, что горизонтальный перенос генов у прокариот — не редкое исключение, а общее правило) и о древовидных составляющих эволюционных траекторий. Он отмечает, что инструментарий и концепции для анализа соотношения древовидных и сетевидных процессов в эволюции еще предстоит разработать.
Весьма интересным представляется привлечение геномных данных к реконструкции симбиогенеза, в результате которого появились эукариоты, и попыткам найти корень «эукариотического древа». Сам процесс эукариогенеза замечательно иллюстрирует роль случая в эволюции (выбор партнеров при захвате археей протомитохондриального эндосимбионта произошел случайно). Кунин описал цепь событий, которые были инициированы симбиогенезом, и для некоторых из них предложил оригинальные объяснения. Это гипотеза «храповика», перетаскивающего гены симбионта в геном хозяина; гипотеза появления структур (ядерной мембраны) и процессов (вырезание интронов), характерных для эукариотической клетки, в результате защиты от вторжения генов симбионта. Возникновение рекомбинации и полового размножения у эукариот заместило утраченную (или, по крайней мере, сильно ограниченную) способность к горизонтальному переносу генов, который обеспечивал генетическое разнообразие, необходимое для эволюции.
Не менее интересна гипотеза о росте сложности генома не в результате адаптации, а в результате «недосмотра» очищающего отбора. На огромных популяциях прокариот такой отбор успевает оптимизировать геномы и убрать из них слабовредные мутации и некодирующий «мусор», поэтому в геномах прокариот «существенная» информация размещена гораздо плотнее. Популяции эукариот имеют более низкую численность, и очищающий отбор в них слабее. Сохранение элемента в геноме может быть связано с тем, что этот элемент адаптивен, необходим для выживания, а может объясняться и тем, что слабый очищающий отбор не успевает устранить неадаптивные предковые особенности и в их геномах большая доля последовательностей отбору безразлична и потому неконсервативна.
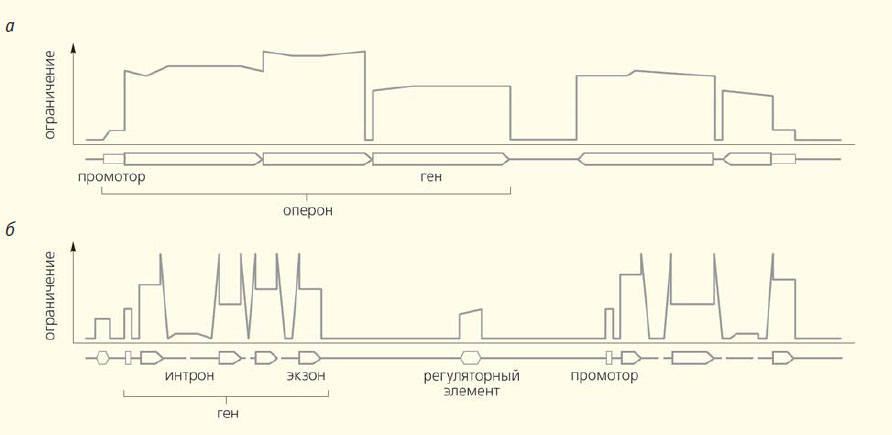
Схема распределения эволюционных ограничений в геномах прокариот (а) и эукариот (б). В геномах эукариот есть протяженные участки, «безразличные» отбору, последовательности нуклеотидов в них высокоизменчивы (см. рис. 3-7, с. 82)
В предложенном Куниным описании не учитывается, что эукариоты возникли и жили в среде, значительно отличающейся от той, в которой проходило становление первичных прокариотических клеток. Прокариоты были не только организмами, из геномов которых компоновались ДНК-последовательности эукариот, но и частью среды (пищей, конкурентами, паразитами, хищниками и т. д.), к которой эукариотам пришлось адаптироваться. Именно присутствие прокариот в среде могло задавать некоторые адаптивные свойства эукариотической клетки (например, размер), реализованные с помощью описанных автором молекулярных механизмов с сильной стохастической составляющей. Можно было бы порассуждать о взаимоотношениях эволюции эукариот с миром прокариот за рамками симбиогенеза, но ведь автор всеохватывающего труда и не обещал.
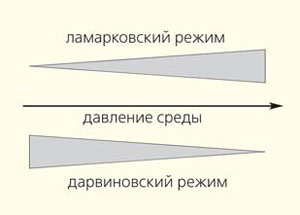
Переход от дарвиновского к (квази)ламарковскому режиму эволюции в зависимости от давления среды (см. рис. 9-3, с. 312)
Принципиально важны для понимания эволюции утверждение, что «способность эволюционировать тоже эволюционирует» (с. 450), и предложенное в книге многофакторное представление эволюционного процесса. В зависимости от давления среды и размера популяции выделены три эволюционных режима. В популяциях малого размера случайные мутации фиксируются случайно, в результате дрейфа (райтовский режим). В больших популяциях факторы среды влияют на отбор. При слабом ее давлении эволюция идет в дарвиновском режиме, при сильном — в (квази)ламарковском. В качестве немногочисленных примеров последнего приведены горизонтальный перенос генов, CRISP-система антивирусного иммунитета бактерий и стресс-индуцированный мутагенез. Приставка «квази» означает, что механизм не чисто ламарковский (он кратко и емко описан в главе 9), а такой, в котором стимул из внешней среды используется для изменения генома, а возникающая модификация адаптивна (напрямую влияет на адаптацию к фактору, вызвавшему изменение) и наследуется потомками клетки.
Происхождение жизни
Сравнение геномов архей и бактерий позволило реконструировать генный репертуар последнего общего предка клеточных форм жизни (Last Universal Common Ancestor, LUCA). В пользу существования LUCA свидетельствуют следующие аргументы: сходство систем трансляции всех организмов — общий генетический код (соответствие нуклеотидных триплетов кодируемым ими аминокислотам); сходная структура рибосом (в их состав входит три универсально консервативных рРНК и несколько десятков белков, из которых 20 универсальны); универсальные компоненты системы трансляции (она включает 30 тРНК, несколько ферментов и других белков). Кроме того, консервативны гены трех субъединиц РНК-полимеразы — молекулярной машины, синтезирующей РНК на матрице ДНК. Помимо этого набора LUCA имел гены, кодирующие ферменты основного энергетического обмена, биосинтеза аминокислот, субъединицы протонной АТФазы и SRP-частиц (частиц узнавания сигналов). Однако в список не попадают ключевые компоненты репликации ДНК и основные ферменты биогенеза мембран, так как они сильно отличаются у бактерий и архей. Это сочетание фактов может объясняться тем, что LUCA был бесклеточным организмом с вирусоподобными РНК-геномами, который реплицировался рибозимами. «Коллекции» РНК-организмов населяли неорганические ячейки и обменивались генетическим материалом. Синтез белка (трансляция) появился позже, синтез ДНК и клеточная мембрана — еще позднее. Кунин отмечает, что одним из центральных моментов появления клеточных форм жизни служит переход от отбора на уровне отдельных генетических элементов к отбору на уровне ансамблей таких элементов. «Важно заметить, что такой переход с одного уровня селекции на другой, по-видимому, лежит в основе многих принципиальных инноваций в эволюции жизни», — пишет Кунин (с. 385), при этом он ссылается на знаменитую книгу Дж. Мейнарда-Смита и Э. Сатмари [4]. Английский эволюционный биолог Мейнард-Смит известен развитием концепции эволюционно стабильной стратегии, которая, будучи принята достаточно большим числом членов популяции, не может быть вытеснена никакой другой стратегией. Его венгерский коллега Сатмари разработал математические модели, показывающие возможность группового отбора в ансамблях репликаторов, самовоспроизводящихся в среде, разделенной на ячейки. Кооперация путем разделения функций, согласно цитируемой Куниным работе Дж. Морриса с соавторами [5], — один из главных факторов эволюции микробных сообществ, и такая кооперация могла осуществляться в доклеточных генетических ансамблях. В этом разделе стоило бы также упомянуть книгу нашего выдающегося физика и кибернетика В. Ф. Турчина о метасистемном переходе и его проявлениях в эволюции мира — от простейших одноклеточных организмов до возникновения мышления, развития науки и культуры [6]. Книга была написана, замечу, еще в 1970 г., правда, издана она только в начале 1990-х.
В примечании к русскому переводу Кунин отмечает, что в своей книге он уделил недостаточно внимания переходу селекции с одного уровня на другой и что в следующем издании (если оно состоится) это будет исправлено. На мой взгляд, отсутствие анализа переходов отбора с одного уровня на другой обедняет рассмотрение эволюции и такое дополнение было бы принципиально важным (возможно, ключевым) дополнением в понимании эволюции сложности.
В разделе о происхождении жизни Кунин рассматривает различные модели возникновения клеточных форм жизни. Роль вирусов в доклеточной эволюции и в клеточном мире в изложении автора оказывается гораздо весомее привычной. Он также приводит свидетельства в пользу мира РНК, рассказывает о происхождении репликации и трансляции, о природе и возникновении генетического кода, об эволюции белковых доменов, отмечая, что многие из предложенных сценариев остаются гипотетичными и не имеют достаточных экспериментальных подтверждений.
Стоит остановиться на красивом примере, иллюстрирующем возможности тотального сравнения структуры белков, — на сценарии предполагаемого происхождения АТФ-синтазы [7]. Эта своего рода молекулярная динамо-машина превращает разность потенциалов на мембране (трансмембранную разность концентраций протонов или ионов натрия) в механическую работу (вращение ротора молекулярной машины), при которой из АДФ и фосфата синтезируется АТФ. Поиск белков, сходных по структуре с компонентами АТФ-синтазы, позволил предположить, что ее статор (заякоренные в мембране части) происходит из мембранного канала, а ротор — из хеликазы (молекулярной машины, умеющей вращаться). В современных клетках хеликазы участвуют в важных клеточных процессах (репликации, репарации, транскрипции, трансляции, сплайсинге и др.). Они расплетают двунитевые участки ДНК или РНК, вращаясь вокруг расплетаемых молекул и обеспечивая свою работу гидролизом АТФ или ГТФ. Хеликазоподобная конструкция предположительно послужила основой транслоказы, объединяясь с мембранным каналом во время транслокации РНК или белка через древние мембраны. Затем, с небольшими модификациями, транслоказа могла быть использована для выполнения другой функции, тоже требовавшей вращения. При этом временная конструкция из трех компонентов стала постоянной — вместо транслоцируемых молекул статор и ротор АТФ-синтазы соединены белковыми субъединицами.
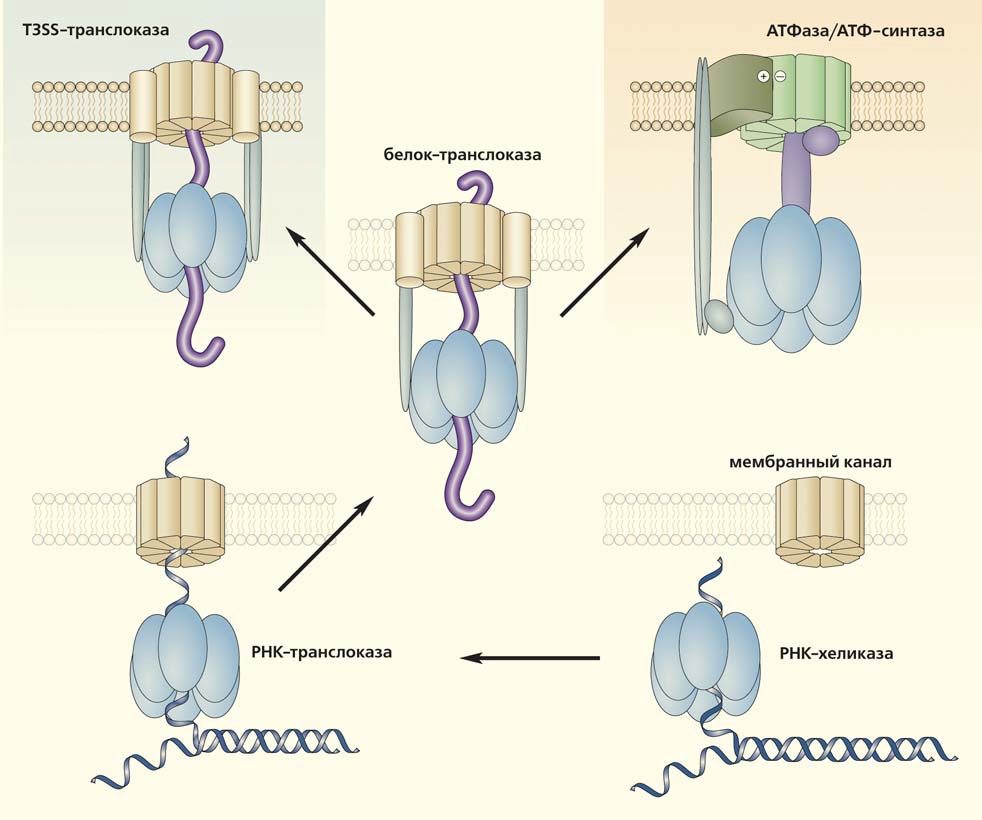
Модель эволюции молекулярных моторов: происхождение АТФ-синтазы и бактериальных T3SS-транслоказ из транслоказ, возникших при переходе временного объединения хеликаз с мембранными каналами (при транслокации РНК и белков) в постоянное (см. рис. 11-3, с. 388). Объяснение в тексте
В целом прекрасное изложение гипотез происхождения жизни и их пока далекого от исчерпывающего эмпирического обоснования приводит на память легендарный совет Н. В. Тимофеева-Ресовского обращаться за детальными описаниями происхождения жизни к А. И. Опарину, поскольку сам он «был тогда еще маленький» и не помнит. Теперь становится ясно, что за этими описаниями надо обращаться к Е. В. Кунину.
Значительный интерес представляют разделы «Происхождение жизни с точки зрения химии и геохимии» и «Радикальная альтернатива: космология вечной инфляции, переход от случайности к биологической эволюции в истории жизни и переоценка роли крайне редких событий в эволюции». Содержание разделов следует из их названия. По поводу последнего автор пишет в примечании, что изложенные в этом разделе идеи могут показаться читателям противоречащими здравому смыслу и представлениям о научном методе, но тем не менее считает их важными (хотя и не главными) для обсуждения новых представлений обо всем эволюционном процессе. Понять этот раздел до такой степени, чтобы кратко пересказать, удалось Г. Ю. Любарскому, к рецензии которого могут обратиться те, у кого возникнет желание ознакомиться с более простым, чем в книге, изложением связи происхождения жизни и современной космологии.
Среди прочего Кунин отмечает, что появлению первых репликаторов, ознаменовавшему начало эволюции путем естественного отбора (и дрейфа) предшествовала череда шагов химической эволюции, на которых биологические эволюционные механизмы еще не действовали. В приложении II он дает грубую оценку вероятности появления первых репликаторов — она оказывается очень низкой. Однако здесь можно не согласиться с автором. Представление, что эволюция путем естественного отбора (и дрейфа) началось только с появлением знакомых нам форм репликаторов вполне дискуссионно. Мы просто пока не знаем, какими могли быть репликаторы до появления нуклеиновых кислот. Гипотеза донуклеиновых репликаторов повышает вероятность известного нам результата химической эволюции. Можно попытаться рассматривать не мономеры и олигомеры — предшественники нуклеиновых кислот, а процессы, субстраты которых могли потом заместиться нуклеиновыми кислотами. Впрочем, эмпирических оснований для таких фантазий пока существенно меньше, чем для гипотез, высказанных автором книги.
На пути к третьему синтезу
«Логика случая» убедительно обосновывает необходимость третьего эволюционного синтеза, описывает полученные в экспериментах факты и их интерпретации, приведшие к смене парадигм, и ясно демонстрирует логику и возможности современного научного исследования и очерчивает широкую исследовательскую программу в области эволюционной биологии. «Понимание простых принципов эволюционной теории незаменимо для самых актуальных и практически важных исследований в современной биологии», — пишет Кунин. В том числе и для медицинских приложений геномных исследований, поскольку, как неоднократно подчеркивается в книге, геном не детерминирует фенотип и связь между ними достаточно сложна, что еще предстоит изучить и понять.
Даже если часть идей, высказанных в этой книге, будет со временем опровергнута, обойти труд Кунина в грядущем синтезе невозможно. Можно высказать замечания по содержанию книги (помимо немногих приведенных здесь, см. рецензию Гельфанда) и оформлению (например, понимать текст было бы легче, если бы книга содержала список сокращений), но сам выход книги такого масштаба — большое событие в российском интеллектуальном пространстве.
Литература
1. Koonin E. V. The Logic of Chance: The Nature and Origin of Biologicval Evolution. New Jersey, 2011.
2. Waddington C. H. The Strategy of the Genes. L., 1957.
3. Wilson D. S., Wilson E. O. Evolution «for the good of the group» // American Scientist. 2008. V. 96. № 5. P. 380–389.
4. Maynard Smith J., Szathmáry E. The Major Transitions in Evolution. Oxford, 1995.
5. Morris J. J., Lenski R. E., Zinser E. R. The Black Queen Hypothesis: evolution of dependencies through adaptive gene loss // Mbio. 2012. V. 3. № 2. e00036-12.
6. Турчин В. Ф. Феномен науки: кибернетический подход к эволюции. М., 1993.
7. Mulkidjanian A. Y., Makarova K. S., Galperin M. Y., Koonin E. V. Inventing the dynamo machine: the evolution of the F-type and V-type ATPases // Nature Rev. Microbiol. 2007. № 5. P. 892–899.





Адаптивный ландшафт Райта (в рецензируемой книге см. рис. 1-2, с. 27) и эпигенетический ландшафт Уоддингтона [2]