Обнаружено латеральное торможение в обонятельных рецепторах насекомых

Давно известно, что хеморецепторные нейроны (нейроны, чувствительные к восприятию химических стимулов) объединены в небольшие комплексы, такие как вкусовые сосочки у млекопитающих или сенсиллы у насекомых. Как правило, нейроны одного комплекса чувствительны к разным веществам. Исследование обонятельных сенсилл у дрозофилы показало, что первичный анализ смеси запахов может осуществляться уже на уровне рецепторных нейронов благодаря механизму латерального торможения. Более того, это торможение, по-видимому, осуществляется без участия синапсов.
Химические органы чувств часто представляют собой группы рецепторных клеток. У млекопитающих, например, это вкусовые сосочки на языке. У насекомых это сенсиллы — видоизмененные участки покровов тела, к которым подходят окончания рецепторных нейронов.
Рецепторные (чувствительные) нейроны — это нервные клетки, воспринимающие определенные виды раздражений. Рецепторные нейроны принято делить на группы: механо-, термо-, баро-, хемо-, фоторецепторы и так далее. Хеморецепторы, в свою очередь, подразделяются на обонятельные и вкусовые.
Наружные части сенсилл насекомых чрезвычайно многообразны. Они могут иметь вид длинного волоска, щетинки, конуса, купола или даже ямки. В составе механочувствительных сенсилл всегда расположена одна рецепторная клетка. Напротив, обонятельные и вкусовые сенсиллы содержат по нескольку рецепторных клеток. Как правило, рецепторные нейроны в составе одной сенсиллы чувствительны к разным веществам и их аксоны проецируются в совершенно различные области мозга. Поскольку до последнего времени считалось, что эти рецепторы работают независимо, то такая организация сенсилл оставалась непонятной. Почему бы не поместить в сенсиллу рецепторы, чувствительные к одному веществу?
Американские ученые из Йельского университета и Монелловского центра изучения обоняния и вкуса (Monell Chemical Senses Center) в Филадельфии подробно исследовали работу обонятельных сенсилл у дрозофилы, чтобы разобраться, почему разные рецепторные нейроны собраны в одной сенсилле. Для этого они выбрали определенный тип сенсилл, расположенный на антеннах мухи, который включает в себя два рецепторных нейрона (рис. 1а).
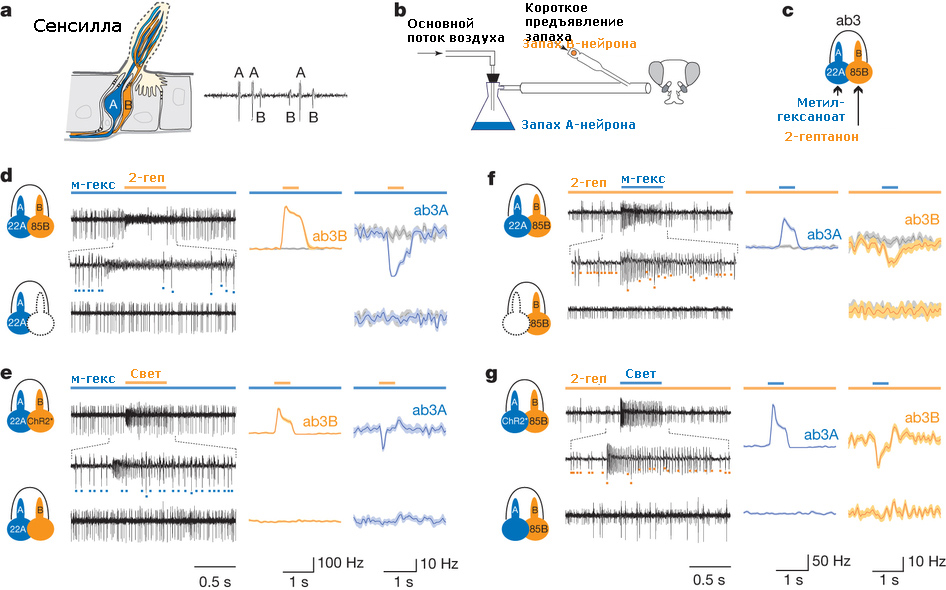
Рис. 1. Латеральное торможение обонятельных нейронов. а. Обонятельная сенсилла, содержащая два рецепторных нейрона — А и В. Справа приведен пример записи активности двух этих нейронов, большие импульсы — нейрон А, маленькие — нейрон В. b. Схема раздражения двумя запахами. c. Сенсилла ab3, для которой показано, что клеточный рецептор OR22A возбуждается метилгексаноатом, а рецептор OR85B — 2-гептаноном. d. Пример возбуждения нейрона ab3А при длительном предъявлении метилгексаноата (м-гекс, большие импульсы) и нейрона ab3B при коротком предъявлении 2-гептанона (2-геп, маленькие импульсы). На записи импульсной активности видно, что ответ ab3А тормозится во время возбуждения ab3B. Справа показаны усредненные ответы для двух нейронов. Нижняя запись импульсных ответов ab3А получена после генетического выключения нейрона ab3b. e. В нейроне ab3B экспрессировали особый тип пигмента родопсина, поэтому стимуляция светом вызывала импульсную активность ab3B, в результате чего тормозился ответ ab3А на метилгексаноат. Внизу приведен контроль: та же стимуляция, но у нормальных мух. Справа — усредненный ответ. f. Стимуляция метилгексаноатом на фоне 2-гептанона — ситуация, противоположная показанной на d. g. Результат экспрессии родопсина в клетке ab3А — ситуация, обратная показанной на е. Изображение из обсуждаемой статьи в Nature
Напомним, что рецепторные нейроны, как и другие нейроны, передают информацию посредством импульсов, или спайков.
Из-за того что концентрации различных ионов внутри и снаружи клетки различны, на клеточной мембране поддерживается разность потенциалов. У клетки в невозбужденном состоянии суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней (то есть внутренняя часть мембраны заряжена отрицательно относительно внешней); это так называемый потенциал покоя). К мембране нейрона в разных местах подходят, образуя синапсы, многочисленные аксоны от других нейронов. Сигналы, поступающие через эти синапсы, слабо и временно изменяют мембранный потенциал в положительную или отрицательную сторону — то есть они могут деполяризовать мембрану, немного выровняв ее потенциал, или наоборот гиперполяризовать ее, увеличив разницу между зарядом внешней и внутренней ее части (cигналы суммируются между собой в пространстве и времени).
Если деполяризация мембраны аксонного холмика (участка, окружающего аксон) достигает некоего порогового уровня, открываются потенциал-зависимые натриевые ионные каналы, которые избирательно пропускают внутрь клетки ионы натрия. Последние устремляются туда просто по градиенту концентрации, потому что снаружи этих ионов на порядок больше. Поток ионов натрия вызывает еще большее и быстрое изменение мембранного потенциала, и волна деполяризации бежит по мембране аксона: происходит генерация потенциала действия, или нервного импульса. Импульс генерируется по закону «всё или ничего», то есть мембрана клетки либо не отвечает на стимул совсем, либо отвечает с максимально возможной силой. (Подробнее о мембранном потенциале, синапсах и импульсах можно прочесть в заметке Обнаружен еще один способ, с помощью которого нейроны контролируют свою возбудимость, «Элементы», 15.07.2010).
В классической электрофизиологии мембранный потенциал измеряется как разность напряжений между отводящим электродом, непосредственно воткнутым в клетку (тогда методика называется внутриклеточной регистрацией) или подведенным как можно ближе к ней (внеклеточная регистрация), и электродом сравнения, находящимся от этой клетки на некотором удалении.
Внеклеточная регистрация показала, что в обонятельной сенсилле разные нейроны генерируют импульсы разной силы (рис. 1а). Тот рецептор, который генерирует большие спайки, принято условно называть клеткой А, а другой — клеткой В. Как правило, это объясняется тем, что нейрон А просто-напросто больше, чем нейрон В.
Авторы экспериментировали с сенсиллами типа ab3. Нейрон A в этих сенсиллах экспрессирует клеточный рецептор OR22A, чувствительный к веществу метилгексаноату, а нейрон В — клеточный рецептор OR85B, чувствительный к 2‑гептанону. То есть нейроны A и B в этой сенсилле возбуждались от метилгексаноата и 2‑гептанона соответственно.
В ходе эксперимента вначале на сенсиллу подавался один химический раздражитель, а затем, на его фоне, коротко предъявлялось второе вещество (рис. 1b). Обнаружилось, что если ab3 долго стимулировать метилгексаноатом, клетка ab3A генерирует импульсы в течение всей стимуляции (рис. 1d). Однако если на фоне этой стимуляции дать короткий стимул 2‑гептанона, импульсный разряд дает другая клетка, ab3B, а первая клетка как раз на это время замолкает.
Торможение клетки ab3A можно было бы объяснить прямым торможением (то есть гиперполяризацией) рецептора OR22A 2‑гептаноном либо каким-то влиянием клетки ab3B на клетку ab3A. Чтобы разобраться, в чем тут дело, авторы использовали дрозофил, у которых экспрессировали специальный ген клеточной смерти, который выключил функционирование клетки ab3B. После этого тормозный эффект при стимуляции 2‑гептаноном полностью исчез (рис. 1d). Таким образом, торможение ab3A вызывалось именно клеткой ab3B.
Чтобы доказать последнюю гипотезу, авторы применили достаточно изысканный метод. В клетке ab3B экспрессировали особый тип пигмента родопсина, чтобы сделать эту клетку чувствительной к световому стимулу (рис. 1е). Активация ab3B светом также тормозила активность ab3А, вызванную метилгексаноатом. С другой стороны, световой стимул не оказывал никакого влияния на активность ab3А у контрольных мух.
Авторы также проверили, происходит ли торможение в обратном направлении. Для этого они начинали предъявлять 2‑гептанон, а на фоне этой стимуляции коротко давали метилгексаноат. В результате получили знакомую картину: во время предъявления метил гексаноата активность ab3B тормозилась (рис. 1f). Генетическое выключение клетки ab3А и экспрессия в ней родопсина привели к тем же самым результатам, что и манипуляции с клеткой ab3B (рис. 1f, g).
Были проверены другие типы обонятельных сенсилл у дрозофилы, а также обонятельные сенсиллы у комара. Понятно, что в разных случаях использовали для раздражения разные химические вещества, которые специфически раздражали исследуемые рецепторы. Во всех этих экспериментах были получены сходные эффекты, описанные выше. Вообще, у этого феномена есть вполне определенное название, описанное в биологии — латеральное торможение. Об этом феномене говорят тогда, когда две или более нервных клеток взаимосвязаны таким образом, что возбуждение одной вызывает торможение другой. Наиболее изучено латеральное торможение в зрительной системе; показано, что такое торможение в большой мере ответственно за остроту зрения.
В обонятельной системе насекомых на уровне рецепторных нейронов этот феномен обнаружен впервые. Авторы пошли дальше — они задались вопросом, каким образом происходит торможение. Логично было бы предположить, что информация передается классическим путем через синапсы — места контакта между нейронами.
Передача импульсов осуществляется химическим путем с помощью медиаторов или электрическим путем посредством прохождения ионов из одной клетки в другую. При возбуждении нервной клетки медиаторы выделяются через нервное окончание в синаптическую щель (область контакта двух клеток) и активируют рецепторы постсинаптической мембраны. Активация рецепторов вызывает изменение трансмембранных ионных токов, и, в зависимости от медиатора и рецептора, соседний участок мембраны либо деполяризуется (тогда клетка легче возбуждается), либо гиперполяризуется (тогда клетка легче тормозится). Сам медиатор расщепляется в синаптической щели и его компоненты обратно транспортируются через пресинаптическую мембрану.
Для исследования механизма торможения был использован тетанотоксин, блокирующий высвобождение тормозных медиаторов из нервных окончаний. Этот токсин экспрессировали в сенсиллах, после чего провели описанную выше стимуляцию ab3B на фоне стимуляции ab3A. Но тетанотоксин не повлиял на латеральное торможение, и клетка ab3A всё равно затормозилась. Исследователи проводили дополнительные эксперименты, отделяя мушиную антенну от головы и проводя электрические отведения от сенсилл. Торможение сохранилось, и это означало, что латеральное торможение между соседствующими рецепторными нейронами происходит на периферии, а не где-нибудь в центральной нервной системе. Получается, что это торможение происходит без участия химических синапсов. Остаются электрические синапсы, в которых нейроны соединяются высокопроницаемыми контактами с помощью особых двойных пор, которые можно увидеть под электронным микроскопом. Надо заметить, что химические синапсы опытный микроскопист также может также разглядеть на электронных фотографиях. Но никакого намека ни на химические, ни на электрические синапсы не нашли в обонятельных рецепторах дрозофилы.
Авторы предполагают иной механизм латерального торможения — эфаптическую передачу (по-гречески «Ephapsis» означает прикосновение, контакт), основанную на электрическом взаимодействии. Вообще, эфаптическое взаимодействие обнаруживали и в химических синапсах, причем впервые оно было предложено российским физиологом А. Л. Бызовым для объяснения некоторых свойств синаптической передачи в сетчатке (см., напр.: Byzov, Shura-Bura, 1986. Electrical feedback mechanism in the processing of signals in the outer plexiform layer of the retina). Но авторы обсуждаемой статьи предлагают механизм, в котором синапсы вообще не задействованы. Согласно их модели, существует электрический потенциал между двумя областями — лимфой, окружающей дендриты рецепторного нейрона, и гемолимфой, омывающей тела нейронов. Благодаря изменению этого трансэпителиального потенциала меняется величина гиперполяризации и, следовательно, торможения соседнего нейрона. Но эта модель, несомненно, требует дальнейшей проверки.
Какой бы механизм в данном случае ни работал, в приведенном исследовании впервые показано, что латеральное торможение задействовано в обонятельной системе насекомых уже на самом первом уровне восприятия сигнала — рецепторном. Это означает, что анализ сигнала может происходить, во-первых, очень быстро, и, во-вторых, в процессе анализа обостряется чувствительность к специфическому запаху. Если на фоне какого-либо постоянного запаха доносится короткая «вспышка» нового химического вещества, например феромона, сигнал о постоянном запахе вытормаживается, и за счет этого сигнал, оповещающий о присутствии феромона, приобретает большую значимость. Таким образом, насекомое может обнаруживать совершенно мизерные концентрации феромона на большом расстоянии от источника. Поистине, насекомые продолжают поражать нас удивительным совершенством своей организации.
Источник: Chih-Ying Su, Karen Menuz, Johannes Reisert & John R. Carlson. Non-synaptic inhibition between grouped neurons in an olfactory circuit // Nature. 2012. Published online 21.11.2012.
Варвара Веденина
Последние новости


Обонятельные рецепторные нейроны у дрозофилы расположены на антеннах и максиллярных щупиках. На схеме справа показаны дендритные окончания рецепторов, тела рецепторных нейронов (OSN cell bodies) и аксоны. Изображение с сайта jones.kaist.edu